 Relatively small numbers of new neurons are produced in several critical parts of the adult human brain. The most well understand region is the dentate nucleus of the hippocampus where new neurons are incorporated into memory circuits for new information that clarifies old memories. Please see the post on how new neurons remodel memory. Special stem cells in the brain are stimulated by complex signals to create these new neurons. Previous posts showed how even signals from capillary cells are vital to direct stem cells. Now very complex epigenetic signals are found to regulate the production of new neurons.
Relatively small numbers of new neurons are produced in several critical parts of the adult human brain. The most well understand region is the dentate nucleus of the hippocampus where new neurons are incorporated into memory circuits for new information that clarifies old memories. Please see the post on how new neurons remodel memory. Special stem cells in the brain are stimulated by complex signals to create these new neurons. Previous posts showed how even signals from capillary cells are vital to direct stem cells. Now very complex epigenetic signals are found to regulate the production of new neurons.
Epigenetic Signals
 Our understanding of genetic regulation is becoming more and more complex. Less than 2 percent of DNA makes proteins. Possibly up to 40 percent makes RNAs both small and large that do not code for proteins. Many of these RNAs have regulatory functions, both affecting DNA directly, affecting messenger RNAs (the template to make proteins), ribosomes (factories that make proteins) and then the proteins themselves. In the most fundamental chicken and egg problem in biology, DNA codes for proteins and proteins regulate the use of DNA by stimulating and inhibiting sections of the code to be used.
Our understanding of genetic regulation is becoming more and more complex. Less than 2 percent of DNA makes proteins. Possibly up to 40 percent makes RNAs both small and large that do not code for proteins. Many of these RNAs have regulatory functions, both affecting DNA directly, affecting messenger RNAs (the template to make proteins), ribosomes (factories that make proteins) and then the proteins themselves. In the most fundamental chicken and egg problem in biology, DNA codes for proteins and proteins regulate the use of DNA by stimulating and inhibiting sections of the code to be used.
Once DNA is chosen and translated into a messenger RNA molecule, before it can be used to code for a protein in the ribosome, there are many edits of this first pre messenger RNA. Editing of messenger RNA is called “alterative RNA splicing”. This process can edit and produce up to 500 completely different proteins from the original DNA taken up. Previous posts show that RNA alternative splicing is uniquely developed in the human brain and in evolution has been a stimulus for its uniqueness.
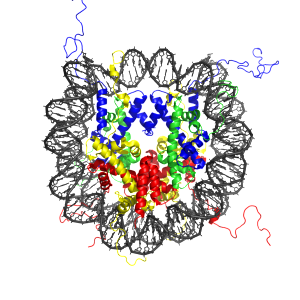
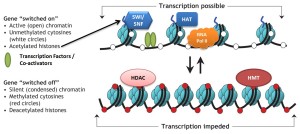
Entirely different levels of regulation exist with specific tags that are placed both directly on the DNA and on proteins (histones) that surround and protect the DNA. These tags on the DNA can either stimulate usage or inhibit usage. There are now more than 40 different tags discovered that operate on the histone molecules that promote or inhibit specific genes. The entire protective material is called chromatin. Recent research shows that at the level of large pieces of chromatin, regulation occurs based on whether chromatin is tightly packed and generally not being used at that moment (heterochromatin) or loosely packed and active (euchromatin). Another factor that determines DNA use is whether it is near the center of the nucleus or the periphery.
 Epigenetics is the name for all regulation of DNA that is not related to specific DNA code and mutations of that code. The impact of epigenetics for stimulating new brain cells is coming into focus. DNA and histone modifications as well as non-coding RNAs are now known to be vital in all stages of producing new brain cells in the adult brain. Likewise, problems in this regulation are correlated with brain disorders. One of the hottest areas of research is newly discovered modification of the code molecule cytosine in DNA (cytosine is one of the four basic letters in the DNA code). Other areas of significance are modifications of messenger RNA different from alternative splicing and alterations on long non-coding RNAs.
Epigenetics is the name for all regulation of DNA that is not related to specific DNA code and mutations of that code. The impact of epigenetics for stimulating new brain cells is coming into focus. DNA and histone modifications as well as non-coding RNAs are now known to be vital in all stages of producing new brain cells in the adult brain. Likewise, problems in this regulation are correlated with brain disorders. One of the hottest areas of research is newly discovered modification of the code molecule cytosine in DNA (cytosine is one of the four basic letters in the DNA code). Other areas of significance are modifications of messenger RNA different from alternative splicing and alterations on long non-coding RNAs.
New epigenetic mechanisms are being discovered each day. There are now more than 40 different tags (modifications on histones and DNA). This post discusses the effect of epigenetic tags on making new brain cells.
New Brain Cells and Epigenetics
 Vast numbers of brain cells are produced in the fetus. In fact, in the ninth month 250,000 new brain cells are minted every minute to make almost a trillion. Then 90 percent are pruned to a mere 80 billion that we live with for most of our life. A small number of new cells are produced in vital brain regions, such as the memory centers in the hippocampus. Recent research (see post) shows that these new brain cells are necessary for new memories in the human brain. These brain cells are produced from stem cells and have simulation from a variety of sources.
Vast numbers of brain cells are produced in the fetus. In fact, in the ninth month 250,000 new brain cells are minted every minute to make almost a trillion. Then 90 percent are pruned to a mere 80 billion that we live with for most of our life. A small number of new cells are produced in vital brain regions, such as the memory centers in the hippocampus. Recent research (see post) shows that these new brain cells are necessary for new memories in the human brain. These brain cells are produced from stem cells and have simulation from a variety of sources.
Many epigenetic mechanisms influence regulation with sets of protein enzymes that place tags (marks) on molecules. Other enzymes take off the marks, but this is sometimes more complex using multiple enzymes. There are another set of protein enzymes that see the mark and respond to it by binding to it and help produce particular effects.
Neural stem cells (NSCs) produce new neurons throughout adult life in the sub granular zone of the dentate nucleus in the hippocampus (SGZ) and the sub ventricular zone of the lateral ventricles (SVZ). Other places can produce cells when there is damage and possibly in the limbic system. It is very hard to find a small amount of new neurons in the vast brain, so this search continues. A previous post showed that all memory and learning is based on these new neurons in the hippocampus becoming active cells in brain circuits.
The cell responds to messages from outside of the stem cells and this produces a series of responses as to the exact place, the amount and the usage of the new neurons.
In the Fetus
 For more details on building of the fetal brain see a previous post. Neurons start with radial glial cells near the ventricles. This starts from epithelium and then more general stem cells appear that produce neurons and glia. These are triggered by epigenetic signals. Later they make specific neuronal stem cells and those that make astrocytes and myelin producing oligodendrocytes. These same stem cells remain as the source of adult neurons.
For more details on building of the fetal brain see a previous post. Neurons start with radial glial cells near the ventricles. This starts from epithelium and then more general stem cells appear that produce neurons and glia. These are triggered by epigenetic signals. Later they make specific neuronal stem cells and those that make astrocytes and myelin producing oligodendrocytes. These same stem cells remain as the source of adult neurons.
These stem cells create the global architecture. In the adult production is very limited. Many of the epigenetic mechanisms occur in both the fetus and in the smaller production in the adult.
Placing Methyl Groups on DNA
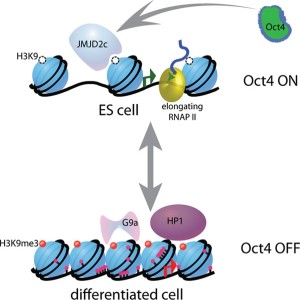 One of the best-known epigenetic mechanisms is placing of methyl groups on DNA or histones. These are attached to particular nucleotides of the DNA or amino acids on the histone proteins. Methyl groups are often placed on the DNA code molecule (called nucleotides) cytosine. Cytosine’s complementary nucleotide is guanine so they are often called CG. When there are many near each other they are called CpG islands. When these are given methyl tags they tend to silence one of the two genes, from a particular parent—such as silencing all father’s genes of this type so that only mother’s function.
One of the best-known epigenetic mechanisms is placing of methyl groups on DNA or histones. These are attached to particular nucleotides of the DNA or amino acids on the histone proteins. Methyl groups are often placed on the DNA code molecule (called nucleotides) cytosine. Cytosine’s complementary nucleotide is guanine so they are often called CG. When there are many near each other they are called CpG islands. When these are given methyl tags they tend to silence one of the two genes, from a particular parent—such as silencing all father’s genes of this type so that only mother’s function.
But, in neurons methyl tags are placed in different regions. In fact, they occur where there are few CGs.
Multiple DNA methyltransferases (DNMTs) enzymes place methyl groups on cytosines in DNA. Some are used for the traditional CpG regions and others are used in the neurons in other areas (DNMT3A and 3B). These are active in the SVZ regions where new neurons are produced in adults. This particular enzyme appears to place tags on many sites very important to making new neurons.
 There are also interactions with enzymes that place tags on histones. What is being learned is that these enzymes have different activities in different stages of the process of making new neurons. Sometimes they stimulate more cells, other times altering the stem cell to produce other kinds of cells.
There are also interactions with enzymes that place tags on histones. What is being learned is that these enzymes have different activities in different stages of the process of making new neurons. Sometimes they stimulate more cells, other times altering the stem cell to produce other kinds of cells.
Seventy five percent of methyl tags placed on cytosine are CG types and a quarter are cytosine next to other nucleotides (adenosine, thymine or another cytosine). When neurons are maturing, there are many tags of this latter type. This type of tag appears to produce unique subtypes of neurons by interacting with proteins that promote specific DNA regions. Unlike the more common CpG type, these seem to be flexible and part of the specific regulation.
Once the mark is placed, special proteins bind to the site and interpret what will happen next. One particular protein (MBD1 or methyl-CpG binding domain protein 1) stops new neurons in the hippocampus. It blocks another protein from producing a factor that promotes new neurons. One of these stimulates the well known BDNF or brain derived neurotrophic factor that promotes new neurons.
As well as these binding proteins to the tag site, other proteins associated with the tags become transcription factors and stimulate very particular gene networks. Contrary to what was thought until recently, these methyl tags increase diverse signals to other genes. Three are known to regulate the process of making new neurons.
Taking off the Methyl Tag
 Previously it was thought that methyl groups become diluted through production of more cells. But, now it is known that specific enzymes actively remove them and these also have impact on regulation of new brain cells. The process of taking off DNA tags is more complex that putting them on and takes several steps.
Previously it was thought that methyl groups become diluted through production of more cells. But, now it is known that specific enzymes actively remove them and these also have impact on regulation of new brain cells. The process of taking off DNA tags is more complex that putting them on and takes several steps.
Proteins involved in taking off methyl groups (GADD45beta or growth arrest and DNA damage inducible protein 45beta) increase new neurons in the hippocampus. They stimulate BDNF and affect proteins that stimulate gene networks. They increase the numbers of neuronal stem cells and these make more neurons. There are several molecules in this process that accumulate over time and are necessary for the health and maturation of neurons over time. This molecule also interacts with histone tags as part of the process.
 Several molecules that are part of the de methylation process become markers on the DNA themselves. They attract other molecules that become enhancers of particular gene networks with the outcome increased neurons in the hippocampus. These molecules appear almost entirely in the brain and not other tissues. These have multiple other molecules that bind to them and process varied effects in the process.
Several molecules that are part of the de methylation process become markers on the DNA themselves. They attract other molecules that become enhancers of particular gene networks with the outcome increased neurons in the hippocampus. These molecules appear almost entirely in the brain and not other tissues. These have multiple other molecules that bind to them and process varied effects in the process.
One particular protein family (TET – ten eleven translocation) produces de methylation by oxidation of cytosine methylation. The different versions work at specific genes. One type works at start sites and another at inhibition sites. They also work with specific co factors. One type decreases the number of stem cells in the SGZ. Others stimulate gene networks that increase the number of neurons. Several of these appear to work together.
Histone Tags and Production of New Brain Cells
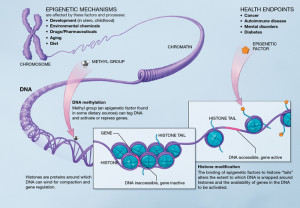 DNA is wrapped around a protein structure made of 8 interconnected histone molecules. There are variant types of the subunits involved. Two copies are used of H2A, H2B, H3 and H4. These are like a spool that the DNA is wound around and each spool has a tail. Marks are placed on tails and these can stop the spool from opening. If the spool doesn’t open the DNA in that section cannot be used.
DNA is wrapped around a protein structure made of 8 interconnected histone molecules. There are variant types of the subunits involved. Two copies are used of H2A, H2B, H3 and H4. These are like a spool that the DNA is wound around and each spool has a tail. Marks are placed on tails and these can stop the spool from opening. If the spool doesn’t open the DNA in that section cannot be used.
It was thought that specific marks had definite effects. But, now it has been found to be more complex. Marks on the tails attract other branching molecules that have multiple different effects. Also there are conversations between the histone tags and the DNA tags. These interactions form a global landscape that in some ways defines the uniqueness of a particular neuron. In particular tags of methyl and acetyl onto lysine amino acids in the histone molecule have very specific effects on making new neurons.
 Methyl marks on histones (onto lysine and arginine amino acids) are more complex because the methyl marks have additional marks on them. They can have multiple additional tags as side chains. Some increase protein production and some do the opposite. There are multiple enzymes to place the tags and to take them off. One of these tags regulates the transition of the stem cell from making neurons to making astrocytes. Another stops this alteration. One type makes the number of stem cells smaller in the SVZ. Some of these marks are related to the particular type of neuron that is to be produced. There are multiple enzymes to remove these tags with many other effects.
Methyl marks on histones (onto lysine and arginine amino acids) are more complex because the methyl marks have additional marks on them. They can have multiple additional tags as side chains. Some increase protein production and some do the opposite. There are multiple enzymes to place the tags and to take them off. One of these tags regulates the transition of the stem cell from making neurons to making astrocytes. Another stops this alteration. One type makes the number of stem cells smaller in the SVZ. Some of these marks are related to the particular type of neuron that is to be produced. There are multiple enzymes to remove these tags with many other effects.
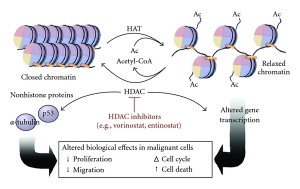
Like methyl groups, acetyls can be both added and reversed by special enzymes. The proteins that take off acetyl groups are called deacetylases and those that work on histones are called histone deacetylases (HDACs). HDACs are very significant in producing neurons. One type decreases the number of very young neurons (neuroblasts) in the migratory stream. This rostral migratory stream is the way new neurons are produced in the SVZ and then travel to the olfactory region where they are incorporated into circuits. This particular HDAC decreases interneurons in the olfactory region. A similar effect also occurs in the cortex during fetus brain development.
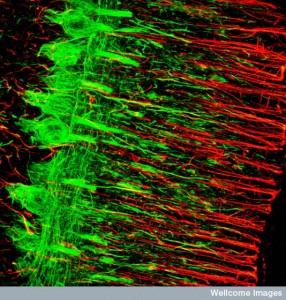 There are now 18 HDACs known that operate in very specific regions to make new brain cells. HDAC2 increases stem cells into neurons. HDAC1 is mainly in glial cells. Another type influences growth of neurons after they are produced. HDACs processes are related to many diseases but there are still a lot not known. One HDAC inhibitor stops new brain cells in one region but increases it in another.
There are now 18 HDACs known that operate in very specific regions to make new brain cells. HDAC2 increases stem cells into neurons. HDAC1 is mainly in glial cells. Another type influences growth of neurons after they are produced. HDACs processes are related to many diseases but there are still a lot not known. One HDAC inhibitor stops new brain cells in one region but increases it in another.
Depakote, a medication used for seizures and for bipolar disorder, is an HDAC inhibitor. It increases stem cells to move in the direction of neurons, but stops the development of new astrocytes and oligodendrocytes. Clearly, there is a lot more to be learned about these markings.
Brain Disorders Related to Epigenetics of New Brain Cells
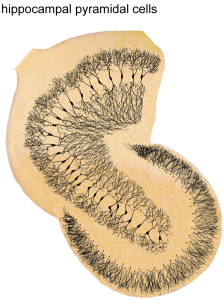
Recent research shows that in depression, stress and other illnesses there are less new brain cells produced. Treatment starts their production again. Alterations in epigenetics that influence this process of new brain cells can produce many brain diseases.
Degenerative brain diseases such as Alzheimer’s, Parkinson’s, (PD) and Huntington’s have impairment in the process of making new neurons. Decreased dopamine in Parkinson’s decreases stem cells in SGZ. PD has less new neurons in hippocampus. A particular molecule α-synuclein is involved in synapses and stimulates new neurons and new connections. But, it is intensely regulated. High levels are found in PD causing death of neurons. Methyl tags affect the levels of the gene making α-synuclein. The special enzyme already mentioned that takes off methyl groups (DNMT1) is very involved in many of the genes related to PD. Another key protein and gene is regulated by the tags that are affected by a microRNA miR-205 (see post on micro RNA in psychiatry). This micro RNA is decreased in PD.
 Alzheimer’s is associated with specific genes in early onset type (PSEN) and Apo lipoprotein E (APOE). New neuron production is affected in both. There is global decrease of tags in the hippocampus cells DNA and histones. These tags related both to acetyl and methyl groups are associated with particular risk genes for disease.
Alzheimer’s is associated with specific genes in early onset type (PSEN) and Apo lipoprotein E (APOE). New neuron production is affected in both. There is global decrease of tags in the hippocampus cells DNA and histones. These tags related both to acetyl and methyl groups are associated with particular risk genes for disease.
Huntington’s disease (see post on why there is no cure for Huntington’s) has decreased new neurons. Alterations in methylation of genes has been shown to be related as well by promoting longer repeats of specific DNA sequences that are the hallmark of the disease.
Decreased neurons in the hippocampus have been found in depression. Treatment success with both antidepressants and therapies correlates with reversing this trend. As depression is better there is increased new neurons in the hippocampus. BNDF decrease associates with depression and increase with cure. Epigenetic factors regulate BDNF as mentioned already above. Another post showed the relationship of specific microRNAs and psychiatric illness—particularly miR-132. There is less direct evidence in schizophrenia, but it does have decreased new neurons in dentate gyrus of the hippocampus. One study showed changes in tags on histones and DNA for genes related to synapses and BDNF.
RNA Signals
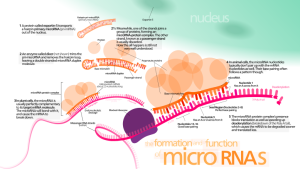
Another area of research is just beginning. It has been recently noted that large areas of what was referred to as “junk” DNA is not junk at all but rather large non coding RNAs. These large RNAs have dramatic epigenetic effects and can silence or affect an entire region of DNA. The smallest of the RNAs, microRNAs, are also noted to be extremely important in many processes.
Epigenetic Signals Regulate New Brain Cells
 Research into epigenetic mechanisms is finding a deluge of different mechanisms and levels of signaling. It is hard to imaging how this is all coordinated and directed. In histones alone, there has been an increase from several tags known a couple of years ago to more than forty currently. Each have multiple placements and effects.
Research into epigenetic mechanisms is finding a deluge of different mechanisms and levels of signaling. It is hard to imaging how this is all coordinated and directed. In histones alone, there has been an increase from several tags known a couple of years ago to more than forty currently. Each have multiple placements and effects.
Directions for these processes must include signals from all of the thousands of microRNAs and large RNAs that do not make proteins. Many proteins are, also, signals for DNA. Tags are used on DNA, histones, on RNAs and on proteins. Multiple tags are placed one on top of another with branching structures increasing complexity exponentially. Three dimensional structures of DNA and chromatin have great influence.
The complexity is astounding and is increasing every day. Yet, all of this is coordinated and all of it responds to mental processes. Is it mind in nature that directs these process?