 When seeking the origin of cellular life, most scientists first try to create a spontaneous fatty membrane that could, theoretically, surround a cell. In fact, real cellular membranes are anything but spontaneous and simple. Membranes are made of extremely complex lipids, of which there are a vast amount of different versions, each with different mechanical and functional properties. Each type has different chemical and structural characteristics and many different functions. Very complex manufacturing centers build these many different lipids, transport them, and signal constantly between compartments for quality control. Each of the thousands of different lipids has different chemical and structural characteristics used for specific functions.
When seeking the origin of cellular life, most scientists first try to create a spontaneous fatty membrane that could, theoretically, surround a cell. In fact, real cellular membranes are anything but spontaneous and simple. Membranes are made of extremely complex lipids, of which there are a vast amount of different versions, each with different mechanical and functional properties. Each type has different chemical and structural characteristics and many different functions. Very complex manufacturing centers build these many different lipids, transport them, and signal constantly between compartments for quality control. Each of the thousands of different lipids has different chemical and structural characteristics used for specific functions.
 Lipids are rich in energy so they are useful for many reactions. They don’t dissolve in water so they are great for barriers in the architecture of the cell. They can, also, protect cargo for transport in round sacs (vesicles), as well as viruses.
Lipids are rich in energy so they are useful for many reactions. They don’t dissolve in water so they are great for barriers in the architecture of the cell. They can, also, protect cargo for transport in round sacs (vesicles), as well as viruses.
While proteins are manufactured through enormously complex genetic machinery (see post Genetic Complexity), lipids are manufactured basically in the large factories of the endoplasmic reticulum ER and refined in the equally complex Golgi apparatus. The ER and the Golgi are, themselves, networks of membranes of many different types, with protein machinery imbedded in different compartments. The transport pipeline of lipids between the ER, the Golgi and the cellular membrane determines much of the function of the cell.
Three Basic Types of Lipids
Lipids provide many cellular functions including storing energy and maintaining supplies of fuel. They have many high-energy bonds of carbon and hydrogen. Many are used as signaling molecules. The most widely known use is to provide the structure of all membranes in the cells, which determine the compartments and functions of all the cell’s organelles.
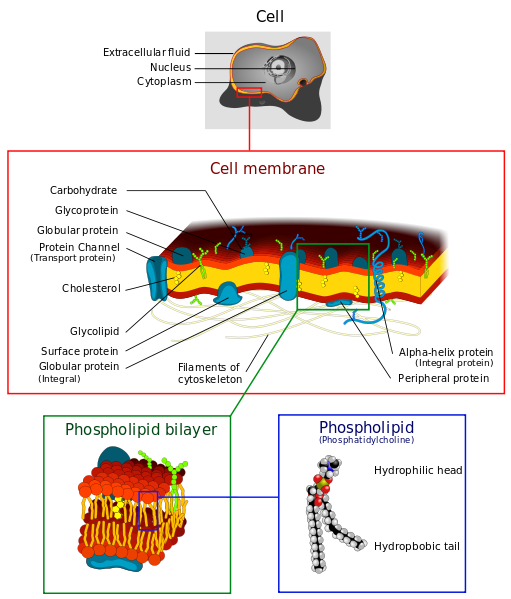
Phospholipids
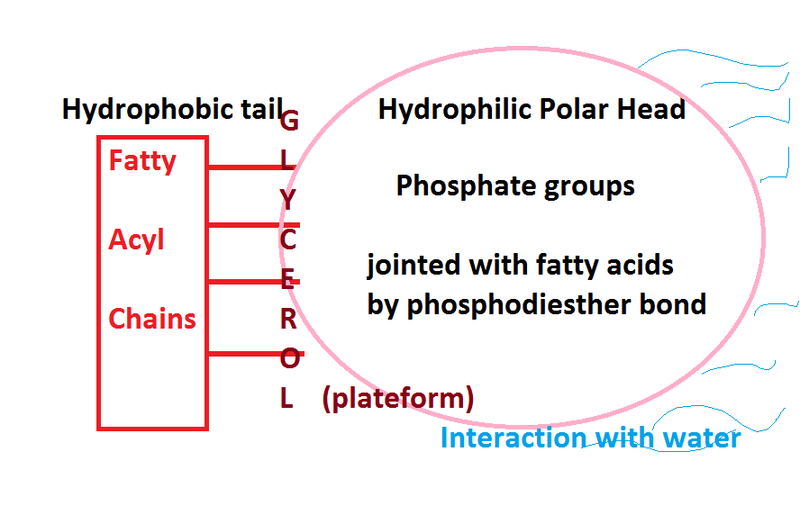
Phospholipids have two chains of carbon and hydrogen called fatty acids, a phosphate molecule (a phosphorus with four oxygen) and a section of glycerol (which is a sugar alcohol). The long tails of fatty acids do not like water (hydrophobic) while their heads made of the phosphates do like water (hydrophilic). Putting two back to back creates a space in the middle with two phobic regions that don’t like water. The outside of the membrane, both inside the cell and outside the cell, have a region that connects well with the water there. This makes double phospholipid layers ideal for membranes.
Glycolipids
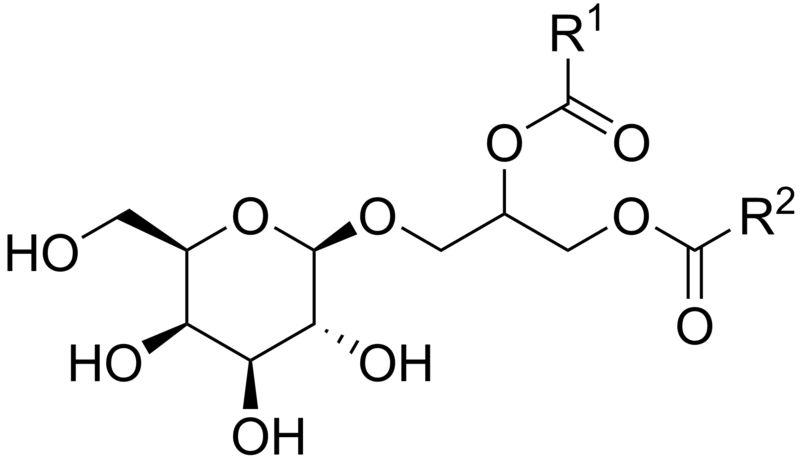 There are a wide variety of lipids that have attached sugars, called glycolipids. A simple version, cerebroside, has only one sugar molecule. There are many very complex versions, such as gangliosides in neurons that have 7 different sugars. The sugar molecule is, always, on the outside of the membrane and is used as a receptor to recognize other cells. Glycolipids, like phospholipids, can also store energy.
There are a wide variety of lipids that have attached sugars, called glycolipids. A simple version, cerebroside, has only one sugar molecule. There are many very complex versions, such as gangliosides in neurons that have 7 different sugars. The sugar molecule is, always, on the outside of the membrane and is used as a receptor to recognize other cells. Glycolipids, like phospholipids, can also store energy.
Cholesterol
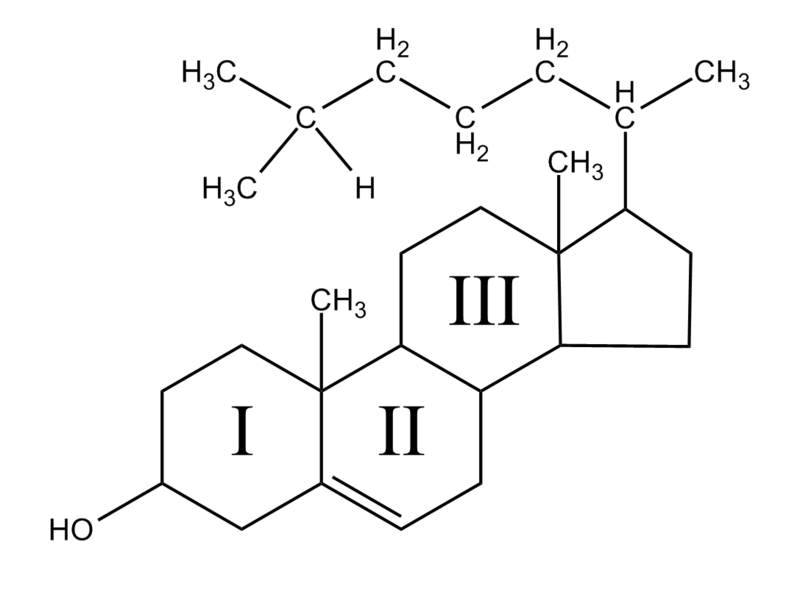
The third type are the sterols, the most famous of which is cholesterol. Cholesterol is well known to cause clogged arteries; in fact, cholesterol has a wide range of types and uses. (A future post will discuss cholesterol’s critical roles in the brain). This lipid has chemical rings made of carbon and hydrogen that have unique properties. The rings are attached to another group of oxygen and hydrogen called hydroxyl. Because of the four rings and the hydroxyl, the molecule is both hydrophilic and hydrophobic. Being partly hydrophobic, it has to be carried by transport proteins in the blood (lipoproteins). One of the many uses of cholesterol is to make membranes. It, also, is the basis of manufacturing steroids, like testosterone and estrogen.
Oil and Water
 Everyone has experienced the separation and mixing of oil and water in salad dressing. Because of the different chemical properties, oils like to stay together and away from water. Water loving chemicals are polar, that is they have subtle differences in positive and negative charges in two different sides of the one molecule. Lipid loving molecules have no charge difference.
Everyone has experienced the separation and mixing of oil and water in salad dressing. Because of the different chemical properties, oils like to stay together and away from water. Water loving chemicals are polar, that is they have subtle differences in positive and negative charges in two different sides of the one molecule. Lipid loving molecules have no charge difference.
Lipids occur naturally in many different forms: fats, vitamins (A, D, E and K), wax, triglycerides, and others. They store energy, signal and make membranes. They are made up of two distinct types of molecules combined—ketoacyl and isoprene—and combinations of these make up many different molecules.
Often lipids are called fats because of one major type of lipid—triglycerides. There are some lipids that must be eaten, but the cell can manufacture most.
Factories for Lipids and Membranes
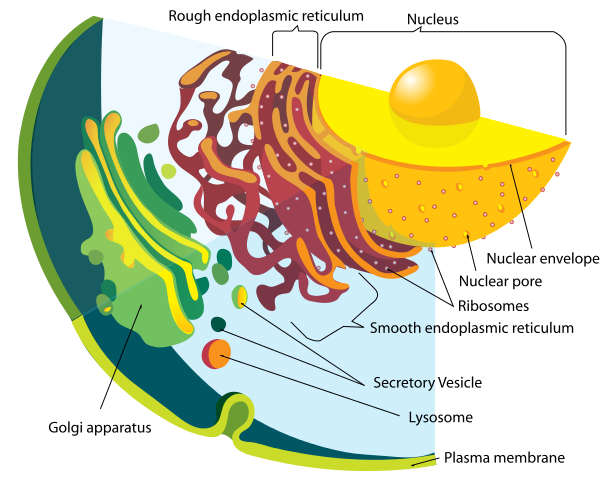
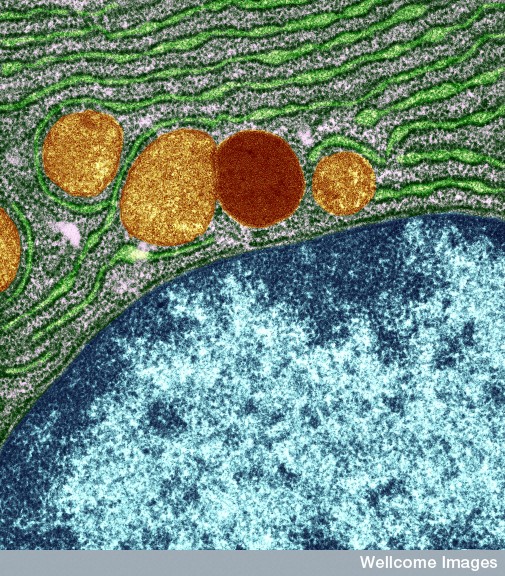 The endoplasmic reticulum sitting right next to the nucleus manufactures proteins and helps them to fold properly. The Golgi next door makes many complex proteins, lipids, and lipoproteins. Previous posts have described the critical communication that occurs in the neuron with mitochondria docking in the ER.
The endoplasmic reticulum sitting right next to the nucleus manufactures proteins and helps them to fold properly. The Golgi next door makes many complex proteins, lipids, and lipoproteins. Previous posts have described the critical communication that occurs in the neuron with mitochondria docking in the ER.
The endoplasmic reticulum (ER) is divided into two types. Adjacent to the nuclear membrane, is the rough ER, called rough because it is studded with many large ribosomes. In the microscope, the many ribosomes give a rough image. This large complex of membrane sheets (cisternae) is right next to the nuclear pores, out of which the messengerRNA travels to meet the ribosomes.
The smooth ER sits contiguous and outside of the rough ER, and does not have ribosomes. It is where lipids are manufactured, altered and maintained. It is the center of membrane and vesicle production. All lipoproteins—combinations of protein and lipid—are manufactured in the smooth ER. It is, also, the center of carbohydrate metabolism, detoxification, and the docking where mitochondria get instructions and molecules from the cell.
 Further outside the ER is the Golgi apparatus, another large set of complex membrane sheets that further modifies all of the lipid and proteins. It provides the tagging for the protein transport and the maintenance of the vesicle factory and transport system.
Further outside the ER is the Golgi apparatus, another large set of complex membrane sheets that further modifies all of the lipid and proteins. It provides the tagging for the protein transport and the maintenance of the vesicle factory and transport system.
The Golgi is the place where proteins are sorted and tagged for their transport to specific parts of the cell. The Golgi packages them into secretory vesicles for use inside the cell or outside (neurotransmitters). The Golgi, also, creates lysosomes, the vesicles used to kill predators. It is the critical factory for the transport of lipids to all parts of the cell. Golgi is the post office that labels each molecule for transport. Vesicles that come from the ER merge with the Golgi and dump its contents, where the molecules are modified before the next transport.
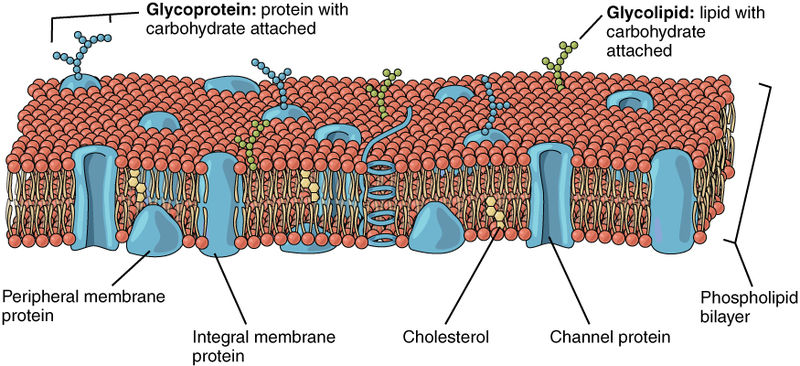
Cellular Membranes
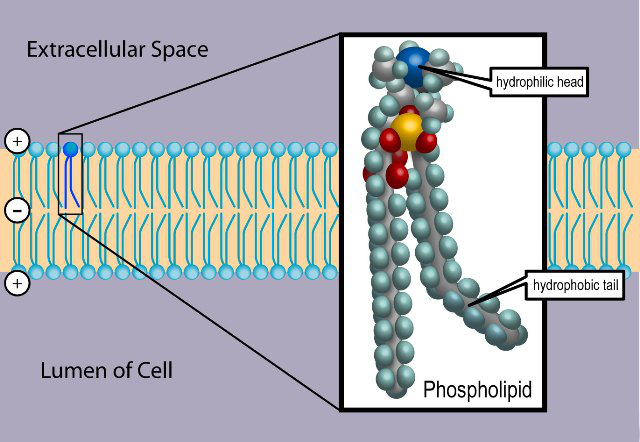
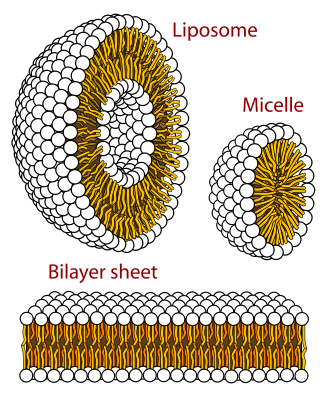
Membranes are formed by two layers of lipids, each of which has a polar side and a non-polar side. The two non-polar sides stick inwards together creating a large lipid region. This lipid-controlled region regulates how molecules move in and out of the cell—water, proteins, nutrients, and others.
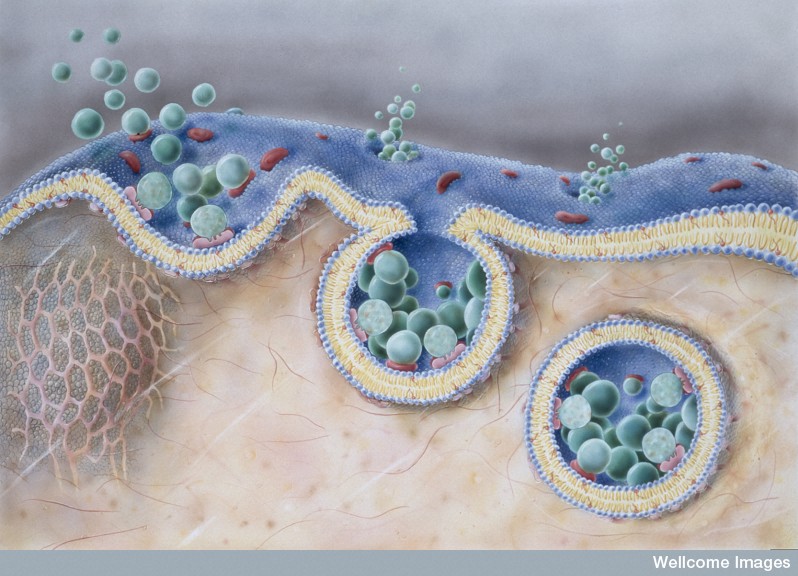
The polar sides stick outwards and inwards connecting with the water based solution. Because of this structure, polar molecules cannot get through the membrane without help, such as a protein channel. This same membrane forms around organelles and vesicles inside of the cells and virus capsules. The virus actually steals some of the cell’s membrane to make its cover, so it can stay whole while it floats in the watery fluids of the extra cellular space.
A simple one layer of lipids would create a compartmental barrier, but wouldn’t have the capacities needed to direct the flow of molecules across or maintain the large protein receptors that lie in the membrane with one part on the outside receiving a signal and other part on the inside triggering a cascade inside the cell.
There Are Many Kinds of Membranes for Different Purposes
 Membrane lipids have a great amount of diversity. Each type of organelle and vesicle uses unique membranes for different functions. The differences in the structures are determined by putting together a large number of different kinds of lipids.
Membrane lipids have a great amount of diversity. Each type of organelle and vesicle uses unique membranes for different functions. The differences in the structures are determined by putting together a large number of different kinds of lipids.
The diversity is in both the types of lipids used and, also, the ways they are put together. Differences include electrical properties, size and thickness, the density of the material and how they are packed together. With these dramatically different membrane structures, there has to be multiple different factories to produce them and different ways that the structures are transported and protected. Also, there are different signals between compartments to produce the different types and a flow of materials between all of the types.
Sensors in the membranes actively send information to the machinery that manufactures the specific lipid layer.
Membrane Characteristics of Phospholipids
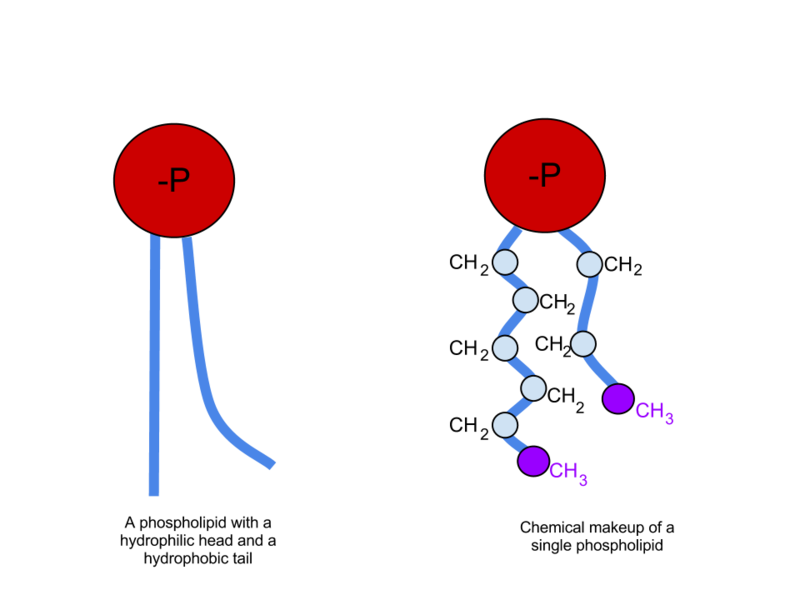
Phospholipids are the most common lipids in membranes. The phosphorus region has many variations and specific characteristics, along with many different side chains. Specific characteristics are related to the length of the side chains and the amount of saturation of the hydrogens attached to oxygen (saturation in fats is a common consideration in health issues).
One very common type uses an oleic unsaturated chain, which decreases the density and increases the flexibility of the membrane. It works best at physiological temperature.
Another type has the shape of a cone with a stressed negative curve. As the chains become more unsaturated, they are less likely to be in a double-layered membrane, but rather a single layer. This type of membrane is good for membranes that fuse and for membranes with large embedded protein molecules. They are less common but are critical for interactions related to electric charge of proteins in the membrane.
Membrane Characteristics of Sphingolipids
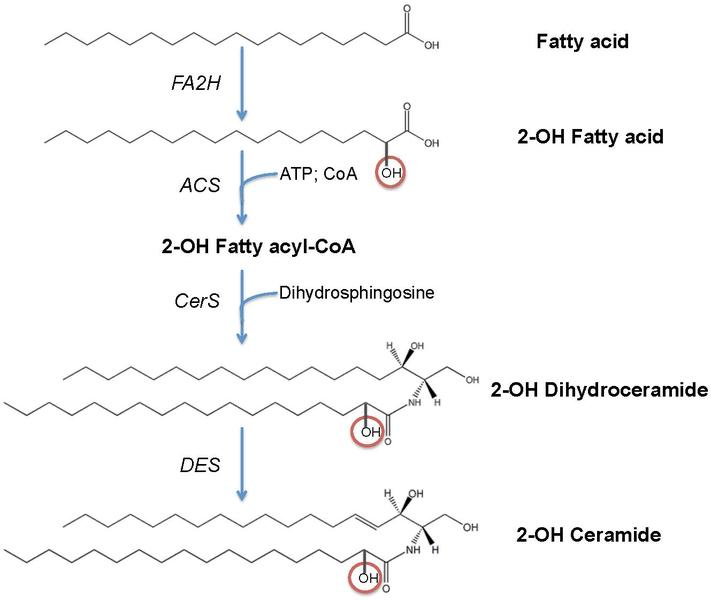
A major sphingolipid is ceramide—hydrophobic, with mostly saturated chains and a serine backbone. It has many different attachments, some of them sugars. It makes a tall very narrow cylinder, which means in can be packed more densely. At physiological temperatures, these double layers are a solid gel and very impenetrable.
If sterols (cholesterol) are attached to ceramides, the membrane becomes more flexible. With sterols, it doesn’t form a gel because the chains can’t pack as tightly.
Strangely, sterols can make some membranes more rigid by making the membranes thicker, but at the same time, less able to have molecules cross the barrier. Therefore, sterols have the unique characteristic of stopping movement across the barrier but, also, making them flexible.
Different Lipids Make Varied Compartment Membranes
 The ER has a very thin bilayer, which is not dense and has no charge – to provide commerce. It uses shorter proteins inside the membrane with 20 amino acids. These characteristics are important for its manufacturing functions.
The ER has a very thin bilayer, which is not dense and has no charge – to provide commerce. It uses shorter proteins inside the membrane with 20 amino acids. These characteristics are important for its manufacturing functions.
The cell’s outer membrane has a thick bilayer, which is dense and has a negative charge on the surface as a barrier. For this it needs longer proteins inside the membrane with 25 amino acids. These characteristics are adapted for barrier function.
The Golgi is the intermediary, which sorts lipids between ER and the outer cellular membrane. The Golgi is polar and has multiple compartments to make each kind of lipid.
Some of the main qualities of lipids for membranes are:
- Short unsaturated fatty acids make it fluid and flexible. Double bonds cause curves and kinks, lower density and stop gels.
- Phospholipids use saturated and unsaturated pairs, which keeps them together despite changes in temperature. Thickness is from chain length and sterols.
 Packing defects occur with unsaturated chains and small heads.
Packing defects occur with unsaturated chains and small heads.- Charge is caused by specific lipids that are like ions (phophatidylserine and phosphoinositides).
- Curvature derives from the lipid shape: small head causes a cone; chains cause a negative curve; equal sized heads causes a cylinder. Very large heads cause an inverted positive curve.
- Cones and cylinders are very strongly regulated.
Lipid Rafts
A special structure, called lipid rafts, floating in the membrane creates a unique organization for a large number of receptors and connections to signaling cascades inside the cell. Lipid rafts are made of many complex molecules — lipids, proteins, lipoproteins, glycoproteins, and glycolipoproteins.
The rafts, also, change the characteristics of the membrane in terms of flexibility; create an organization for large signaling molecules lying in the membrane; and regulate transport of receptors into the membrane and secretion of neurotransmitters.
Lipids Travel Between ER, Golgi and Outer Cell Membrane
 Sterols are rare in the ER but appear in the Golgi and are very common in the outer cell membrane. Ceramide travels from ER to Gogli and the Golgi turns it into sphingolipids, which go to the cell’s outer membrane. Chains are also altered in Golgi and become more saturated in cell membrane.
Sterols are rare in the ER but appear in the Golgi and are very common in the outer cell membrane. Ceramide travels from ER to Gogli and the Golgi turns it into sphingolipids, which go to the cell’s outer membrane. Chains are also altered in Golgi and become more saturated in cell membrane.
The outer cell membrane is dense, rigid and thick to make a good barrier. So, thin membranes from the ER are altered in the Golgi to become dense, rigid and thick by adding sphingolipids and sterols. This alteration is against a gradient and it is difficult for the Golgi to balance the needs of these two very different types of membranes. The Golgi, also, adds specific materials that create a charge and places them in totally different parts of the membranes.
Both the Golgi and the cell membrane have definite charge. In the ER, ions are hidden in a special compartment because the ER t membrane doesn’t have charge.
The Secretory Pathway
 To make proteins that will leave the cell in vesicles, there is the secretory pathway to create and transport vesicles lipids. The series of steps starts in the rough ER where protein is made; it then travels to many different compartments where it is folded and modified, and then sent to the Golgi. The Golgi tags it for secretion and it is placed in a vesicle. The vesicle fuses with the outer cell membrane by a very complex process involving porosomes. Porosomes are very large molecules in the membrane that look like a cup. The vesicles dock there to fuse to release the contents. This complex includes the famous SNARE proteins involved in neurotransmitter function.
To make proteins that will leave the cell in vesicles, there is the secretory pathway to create and transport vesicles lipids. The series of steps starts in the rough ER where protein is made; it then travels to many different compartments where it is folded and modified, and then sent to the Golgi. The Golgi tags it for secretion and it is placed in a vesicle. The vesicle fuses with the outer cell membrane by a very complex process involving porosomes. Porosomes are very large molecules in the membrane that look like a cup. The vesicles dock there to fuse to release the contents. This complex includes the famous SNARE proteins involved in neurotransmitter function.
Sensors Monitor Quality of Varied Necessary Lipids
The cell must transport proteins through totally different environments.
 Composition sensors monitor the membranes for physical characteristics related to specific lipid composition and send signals to alter the manufacturing process and the transport. The ER provides most lipids to other regions of the cell, including creating vesicles and lipids for mitochondria, which they receive by contact with the ER. (see post on mitochondrial function and the ER).
Composition sensors monitor the membranes for physical characteristics related to specific lipid composition and send signals to alter the manufacturing process and the transport. The ER provides most lipids to other regions of the cell, including creating vesicles and lipids for mitochondria, which they receive by contact with the ER. (see post on mitochondrial function and the ER).
The ER responds acutely to any changes in the composition of the lipids being used. An example is very different reactions to saturated and unsaturated side chains.
 The types of saturation and side chains are regulated through special sensors. These sensors are connected to the quality control for proteins called the unfolded protein response (UPR). Proteins in this system sense misfolded proteins, which will alter the lipid synthesis to deal with the stress. Too many fatty acids also trigger UPR.
The types of saturation and side chains are regulated through special sensors. These sensors are connected to the quality control for proteins called the unfolded protein response (UPR). Proteins in this system sense misfolded proteins, which will alter the lipid synthesis to deal with the stress. Too many fatty acids also trigger UPR.
If there are too many sterols the movement of proteins into the ER is blocked. If there are too little sterols then communication with the mitochondria breaks down.
There is an elaborate feedback mechanism in the ER to maintain all of these different forces. Mechanisms include special proteins that sense levels of these molecules. By these mechanisms, sterols and side chains are regulated and sent to all the other organelles in the cell in the appropriate proportions.
There are, also, curvature sensors to make sure that the right lipids are sent. There are very complex signaling cascades that maintain the appropriate curvature for the given organelle.
Lipid Cellular Pathways – The Pipeline
When a lipid has been manufactured in the ER it travels either to make vesicles or it enters a special transport system throughout the cell—the lipid transfer protein pipeline. The pathways for vesicles and the lipid transfer pipeline are completely different.
The pipeline is the source of lipids for mitochondria and other organelles that don’t receive vesicles. A large amount of appropriate lipid is sent to the cell’s outer membrane through the pipeline. One part of the pipeline runs through the Golgi that sends special substances to the membrane, like ceramide and many other very specifically shaped lipids. The transport proteins are essential because without them the lipids will not move rapidly enough through the water filled cytoplasm.
These pipelines are necessary for the great complexity of the different types of lipids that the Golgi must produce and send in specific regions. The pipelines are, also, able to respond to different amounts of the three basic types of lipid classes.
Lipid transfer proteins use a complex procedure. First, a type of lipid binds to a section of the large protein. With the protein changing shape, this binding region then closes so that there is no water in the chamber. Alterations are made to the lipid and then the protein reverses its shape allowing water in and sending the new lipid on its way. The flow of lipids occurs in both directions because these pipelines must maintain an equilibrium between the various types of lipids needed at that moment. There are often mechanical forces in a gradient going against the movement that the protein overcomes.
Many different transport proteins have specific structures for interaction with each different type of lipid. There are larger versions that have two distinct regions to engage two different types of lipids.
Special acceptor organelles work in the pipeline accumulating specific types of lipids for later movement with different mechanisms.
- Metabolic gradients are used, such as converting ceramide to sphingomyelin; in this process only the later is allowed in the special acceptor organelle.
- Thermodynamic characteristics are used with sterols in the ER and different ones in the membrane that allow a specific acceptor to take it in.
- Another technique is the transfer of two specific lipids in opposite directions where the specific characteristic of the lipids and the large protein make the movement occur.
- All of these techniques are, also, used at the same time. Many different mechanisms from the ER to the pipeline proteins to the Golgi occur at the same time.
Signaling Between Compartments for Quality Control
 Sterols and sphingolipids are regulated between the ER and Golgi. There are a large amount of signals sent between these two organelles. Many large enzymes (kinases) respond to the moment-by-moment needs of the all-important cellular membrane. Through these kinase cascades, the ER and the Golgi are constantly communicating with the cellular membrane. (see post, The Remarkable Language of Cells.)
Sterols and sphingolipids are regulated between the ER and Golgi. There are a large amount of signals sent between these two organelles. Many large enzymes (kinases) respond to the moment-by-moment needs of the all-important cellular membrane. Through these kinase cascades, the ER and the Golgi are constantly communicating with the cellular membrane. (see post, The Remarkable Language of Cells.)
Disease
Many diseases can occur with problems in each of these complex pathways, including liver failure in obesity and diabetes. Too many free fatty acids cause liver disease. The accumulation of triglycerides, once thought to be the cause, is now thought to be a result and might actually be protective. Liver cells have problems with too many fatty acids and can’t put them into triglycerides. This causes stress in the ER and then problems in the channels in the membrane. The problem occurs in the ERs inability to regulate the types of lipids.
Amazing Complexity of Cellular Membranes
 There are a very large amount of different lipids used in specific membranes, with many varied properties—shape, packing, energy, and charge. Very complex combinations of multiple lipids are manufactured in the ER and Golgi for each purpose.
There are a very large amount of different lipids used in specific membranes, with many varied properties—shape, packing, energy, and charge. Very complex combinations of multiple lipids are manufactured in the ER and Golgi for each purpose.
The pathways that manufacture and regulate lipids involve two complex transport systems both with special compartments, factories, transport molecules and signals. One is the secretory system to make and manage vesicles. The other is the lipid protein transport pipeline, which sends many lipids throughout the cell using specialized compartments for physical barrier functions related to the cell’s architecture. There are many sensors and complex signals that maintain all of these different functions at the same time, with some tradeoff between them. Many of the cascades and proteins have multiple functions at the same time. They provide for the many different flows, for sensing difficulties, and control the movement of the vesicles.
 The pipeline transports between the early creation of specific lipid molecules and then the production of the altered final complex products that serve very specific roles in the different membranes for vesicles, ER, Golgi, nucleus, nucleolus, mitochondria, lysosomes, exosomes, and the all important complex outer cellular membrane. Also, lipid transport proteins are able to take faulty lipids out of membranes and put in new ones. They, also, have regions that, strangely, do completely different functions such as aiding cell migration and fighting against cancer.
The pipeline transports between the early creation of specific lipid molecules and then the production of the altered final complex products that serve very specific roles in the different membranes for vesicles, ER, Golgi, nucleus, nucleolus, mitochondria, lysosomes, exosomes, and the all important complex outer cellular membrane. Also, lipid transport proteins are able to take faulty lipids out of membranes and put in new ones. They, also, have regions that, strangely, do completely different functions such as aiding cell migration and fighting against cancer.
Most discussions of the complexity of cells is related to the fantastic array of different complex proteins and how they are manufactured through the multi leveled genetic processes. But, equally challenging is the way these proteins manufacture many varied lipids with very specific functions that are totally essential in their own right to the cells existence.
How can anyone think this is a process of lipids randomly coming together? How can trivial randomly created chemical lipid spheres be compared to cellular and organelle membranes? How can anyone not think that complex direction and intelligence is needed for the amazing complexity of cellular membranes?