 How complex is the communication among cells when individual bacteria join together for multi cellularity? Complex bacterial communication has been noted in previous posts. Each individual microbe joins together with comrades for many different activities including complex communication and decision-making. Bacteria in the gut plan ahead, activating genes for maltose enzymes before they encounter it. Recently, it was found that microbes produce electrical wires, made of amino acids that conduct electricity, and supply distance relatives with energy. They signal to change behavior in the colony including building complex cities called biofilms and altering attack modes in animals. Slime molds show intelligent behavior, remembering where it has already travelled. Individual microbes remember how they moved in the past so they can go in a straight line. Microbes send signals that alter their host’s behavior. In a very complex back and forth communication with plants they together gradually build a nitrogen factory.
How complex is the communication among cells when individual bacteria join together for multi cellularity? Complex bacterial communication has been noted in previous posts. Each individual microbe joins together with comrades for many different activities including complex communication and decision-making. Bacteria in the gut plan ahead, activating genes for maltose enzymes before they encounter it. Recently, it was found that microbes produce electrical wires, made of amino acids that conduct electricity, and supply distance relatives with energy. They signal to change behavior in the colony including building complex cities called biofilms and altering attack modes in animals. Slime molds show intelligent behavior, remembering where it has already travelled. Individual microbes remember how they moved in the past so they can go in a straight line. Microbes send signals that alter their host’s behavior. In a very complex back and forth communication with plants they together gradually build a nitrogen factory.
But, combining to become a multi cellular creature is a different level of cooperation. Bacteria have accomplished this multiple times in evolution. On many different occasions the cooperative nature of microbes becomes so central that multi cellular activities become almost a multi cellular organism.
A previous post has described the eukaryote amoeba (eukaryote cells are much more complex than bacteria), who lives as an individual at times and as a multi cellular creature some of the time. Without food, the cells combine to form a slug that travels to a new location. The cells again become individuals, and form a new more complex structure made of a stalk and fruiting body. The fruiting body will fly away either in the air or on an animals foot to a new place to start the colony over. The cells that crawl into the stalk sacrifice themselves by staying behind. It has been shown that stalk cells are more likely to sacrifice themselves for members of their own family.
Bacteria Cells Come Together
 Unlike eukaryote organisms (such as fungus and amoeba) bacteria are a smaller less complex cell, but still have become multicellular several times during evolution. The multi cellular behavior appear to, at least, depend upon aggregation of cells, adhering together as a group of cells, and attachment to an extra cellular matrix. These bacteria, remarkably, transition between single cells and group activity. In E.Coli a single mutation can lead to an alteration into filament growth. Also, stimulation of cell division can reverse a hyphae state back to unicellular.
Unlike eukaryote organisms (such as fungus and amoeba) bacteria are a smaller less complex cell, but still have become multicellular several times during evolution. The multi cellular behavior appear to, at least, depend upon aggregation of cells, adhering together as a group of cells, and attachment to an extra cellular matrix. These bacteria, remarkably, transition between single cells and group activity. In E.Coli a single mutation can lead to an alteration into filament growth. Also, stimulation of cell division can reverse a hyphae state back to unicellular.
When cells aggregate and reproduce as a single unit, individuals within the group are killed in the process or cannot reproduce. These altruistic cells are seen in bacteria, as well as amoeba, more often when they are from the same colony.
Those groups of bacteria that join together from one cell colony (rather than multiple different colonies) have less conflicts socially and can more often differentiate into distinct new cells types that are needed for a multi cellular creature to function. In biofilms, where many different colonies come together, there is more social conflict.
Forms of Bacterial Multi Cellularity
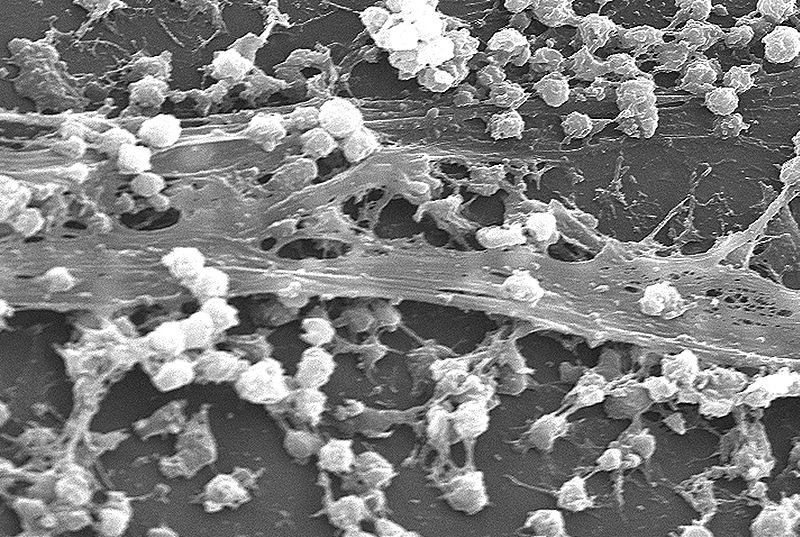 Bacteria live as individuals and some spend time in complex communities. Individual bacteria normally divide into two. They feed independently. Others have left their individual lives and only grow as part of multi cellular creatures.
Bacteria live as individuals and some spend time in complex communities. Individual bacteria normally divide into two. They feed independently. Others have left their individual lives and only grow as part of multi cellular creatures.
Multi cellular organisms are defined as having specific unique shapes, specific patterns of groups of cells, and programmed cell death of individuals to help the larger group. Some employ irreversible differentiation. In real multi cellular organisms specific cells differentiate for specific roles that help the entire colony, not the individual. This means that specific cells can perform important tasks that could not be performed by one cell, such as photosynthesis and nitrogen fixation.
The great advantage of multi cellular life is that the workload can be divided. It was previously noted that bacteria tend to shed their excess DNA to work more efficiently. They can do this if others will pick up functions that otherwise they must do. Forming a multi cellular creature allows work to be divided.
 Biofilms need an extra cellular matrix consisting of polysaccharides, amyloid fibrils, lipids and nucleic acids. The ECM protects against toxins such as antibiotics. At the surface there are enzymes that break up cells nearby as food.
Biofilms need an extra cellular matrix consisting of polysaccharides, amyloid fibrils, lipids and nucleic acids. The ECM protects against toxins such as antibiotics. At the surface there are enzymes that break up cells nearby as food.
Chains are a temporary early state in the creation of biofilms. One mechanism occurs with an altered division process where fission is not complete and a chain is produced as in filamentous cynanobacteria.
When chains occur there is a protein complex linking the cells together. Syncytial filaments have no individual cells, but the contents of many cells together in one large shared space with the many chromosomes. Branching filaments, also, occurr with cell division defects, which can form clusters of cells some of which are apoptotic. But, clustering leads to impaired mobility, increased need for resources, and less buoyancy.
Two benefits of clustering are protection from predators and stress, and cooperative foraging. The much bigger units are hard for the usual predators to eat. Microbes become filamentous with long chains when predators are near, and then again quickly become individual cells again when there are no predators.
Mobile bacteria use these temporary arrangements to avoid predators. Another example is TB, which turns into filaments when it is inside a macrophage to avoid being destroyed. Other microbes suppress full cell division and change into filaments to avoid antibiotics.
These clusters also occur when there is not enough food for individuals to multiply. Incomplete cell separation or clustered growth occurs when there is a need for an enzyme to break down sucrose into glucose and fructose. The smaller mono-saccarides diffuse away from the single cells that are breaking down the sugars and the larger clusters are able to gobble up the glucose and fructose. Also, only the cells in the cluster had access to them.
Biofilms
 In biofilms individual cells are embedded in an extra cellular matrix and multiply while attached. Many different microbes can switch between individual lifestyle and biofilms depending upon circumstances. The cells in the two different environments have extremely different networks of genes activated. In the biofilm they can withstand stress that they could not otherwise, for example, acids, solvents, antibiotics and predators. Biofilms can form at the air-water interface or on solid surfaces. The biofilms that occur at the air water surface, called pellicles, allow cells to have increased oxygen and therefore can grow much faster.
In biofilms individual cells are embedded in an extra cellular matrix and multiply while attached. Many different microbes can switch between individual lifestyle and biofilms depending upon circumstances. The cells in the two different environments have extremely different networks of genes activated. In the biofilm they can withstand stress that they could not otherwise, for example, acids, solvents, antibiotics and predators. Biofilms can form at the air-water interface or on solid surfaces. The biofilms that occur at the air water surface, called pellicles, allow cells to have increased oxygen and therefore can grow much faster.
One type of biofilm forms hills on which spores are created; spores are very tough structures extremely resistant to starvation. To make spores, first cell death occurs in a group of the colony’s cells to extract the nutrients for the spore. This process, called cannibalism can delay the formation of spores.
 In biofilms, cells differentiate into several types to perform specific work. One type of cell forms water channels that are like blood vessels, to transport food and waste products. This differs from vessels in animals and plants, where structures start with one specific new cell that divides to form the vessel.
In biofilms, cells differentiate into several types to perform specific work. One type of cell forms water channels that are like blood vessels, to transport food and waste products. This differs from vessels in animals and plants, where structures start with one specific new cell that divides to form the vessel.
In biofilms, individual cells from different families form aggregate structures, which can lead to conflicts. The biofilm appears to be more a group of colonies that form an aggregate for protection, than specifically differentiated cells. It is more like a school of fish or flock of birds than a multicellular creature. All cells don’t cooperate, but they do on specific defined structures. The cells, however, still maintain definite individuality.
Some biofilms are from only one colony, such as occurs in hospitals. But, in nature more often they are multiple colonies working together. Non-cooperative cheats live in biofilms and hurt the structure, growth and make them more susceptible to antibiotics.
In biofilms, there is complex intercellular signaling, to form of specific structures like spores with the coordination of cannibalism, programmed cell death.
Biofilms have some of the characteristics of multi cellular creatures but they do not have a specific shape and form as multi cellular organisms.
Forming Specific Patterns

In the next level of multi cellular bacterial organisms, coordinated movement of a group of cells forms the basis of the structure forming into specific patterns. These coordinated mobile cells can come from one colony or an aggregate from different cell families.
One example occurs in gram negative myxo bacteria that forage for food as a group of individuals. They move together in a somewhat cooperative activity when feeding. They also put out specific enzymes to break apart cells and large molecules they are after. As they feed with the use of these enzymes, they grow at a specific rate.
During this social feeding they form a rippling pattern with ridges of higher density cells. These ridges move as wave over the prey and make the pattern of enzyme secretion more efficient.
This group forms a much stronger spore, called a fruiting body. Myxobacterial fruiting bodies are much more complex than biofilms.
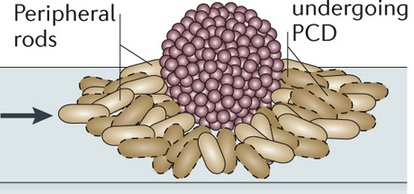 When there is no food, growth stops and a new fruiting body structure forms with a definite shape. This process starts 6 hours after starvation. The fruiting bodies are built over the period of a day and include a hundred thousand cells that are packed together tightly. These fruiting bodies become spores.
When there is no food, growth stops and a new fruiting body structure forms with a definite shape. This process starts 6 hours after starvation. The fruiting bodies are built over the period of a day and include a hundred thousand cells that are packed together tightly. These fruiting bodies become spores.
Specific signals initiate the fruiting body including a protein alarmone and several kinase intercellular signals. At first the cell’s motility changes and new gene networks are triggered. There is an elaborate timing of genetic switches on and off. Elaborate signaling between the cells is necessary. The cells divide into three types for this process. 10% of the cells become spores for the future cells. 30% become rods that are not in the fruiting body. And the remaining 60% die to be used for nutrients. The spore can survive almost anything and can travel to a new region where there is food.
Altruistic behavior occurs when the cells are part of a colony. When they are not part of the colony, cells act in their own individual interest, for example, eating food without contributing to the creation of the enzymes. Cheating, also, occurs in fruiting body formation.
There are mechanisms to determine which cells are not identical and exclude them from the fruiting body. One technique is a surface protein, TraA, used for recognition. Also, specific antibiotics are used to kill non family cells. Some kill all of the cells at the interface of the two groups. In one scenario, the identification of individuals occurs and then some are killed with the antibiotics.
Branching Filaments
 2.5 billion years ago oxygen appeared in the earth’s atmosphere and, at the same time, multicellular cyanobacteria. These bacteria have lost and re-found multicellularity several times. Filamentous cynano bacteria have incomplete separation during cell division. They also differentiate into different cell subtypes for a division of labor. Even though they form distinct cell types, they are always based on one colony with no question of cheaters or conflicts of interest.
2.5 billion years ago oxygen appeared in the earth’s atmosphere and, at the same time, multicellular cyanobacteria. These bacteria have lost and re-found multicellularity several times. Filamentous cynano bacteria have incomplete separation during cell division. They also differentiate into different cell subtypes for a division of labor. Even though they form distinct cell types, they are always based on one colony with no question of cheaters or conflicts of interest.
Individual cells use photosynthesis and nitrogen fixation one process at a time. In the filaments, there are four distinct cells. One uses photosynthesis; one fixes nitrogen; the other two are resting cells and motile cells. These cells are mutually dependent with complementary metabolism. Through communication structures called a heterocyst are formed where intercellular channels send sugar and nitrogen. Once the heterocyst is formed it cannot go back to the previous cell type.
Intercellular channels allow interchange of signals nutrients between cells. Filamentous growth occurs without division through structures called crosswalls, forming a syncytium with many compartments. Transport occurs throughout including plasmids with DNA.
Long Chain Branching
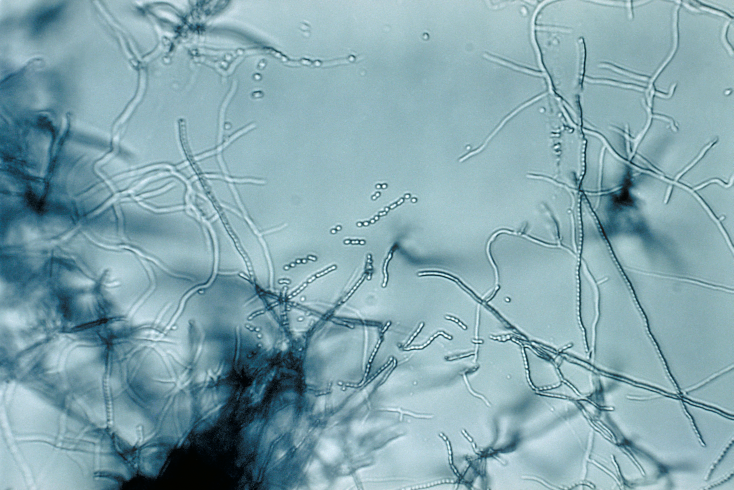 In the next level of complexity, streptomycetes start with one spore cell. These grow as long thin hypha with tip growing and branching. They have many communicating sections separated by crosswalls. There are channels that allow transport of nutrients long distances through the crosswalls.
In the next level of complexity, streptomycetes start with one spore cell. These grow as long thin hypha with tip growing and branching. They have many communicating sections separated by crosswalls. There are channels that allow transport of nutrients long distances through the crosswalls.
When there is a threat, or starvation, some hyphae leave the soil and grow in the air and then alter themselves into a different type of cells to form airborne spores. As the new structure grows in the air, the streptomycete produces an antibiotic, which kills hyphae comrades in the soil and any microbes that might eat the hyphae, but spares the nutrients to be used by the aerobic hyphae. When the aerial hyphae are separated they form chains of waterproof spores that are dispersed to start growth at many other sites.
Most bacteria divide into two with a Z ring at the division site. The long aereal hypha have a unique division process. Ladders of proteins form 100 septa, which then divide into long chains of spore cells. This is a spore specific type of cell division and involves elaborate different signaling pathways and unique proteins. A particular protein family called SALP determines the complexity of this division process. Some species only have one SALP, which means they produce one spore per hypha. Those with up to 14 different SALP proteins produce a large number of spores per hypha.
Individual Bacteria Join Together for Multi Cellularity
 Bacteria understand that there are other members of its colony. They send messages back and forth and help one another. But, exactly, how they can know to become part of a larger organism is not clear. Altruism has been noted in the unicellular creatures because they are more likely to cooperate with these structures and sacrifice their individuality if they are among kin. The knowledge of self versus other begins with individual cells, and continues throughout evolution to the human immune system.
Bacteria understand that there are other members of its colony. They send messages back and forth and help one another. But, exactly, how they can know to become part of a larger organism is not clear. Altruism has been noted in the unicellular creatures because they are more likely to cooperate with these structures and sacrifice their individuality if they are among kin. The knowledge of self versus other begins with individual cells, and continues throughout evolution to the human immune system.
Bacteria demonstrate that individual cells cooperate at many levels—signaling for cooperation and group activity and forming complex structures together. There is a tendency for bacteria to drop excess baggage to allow efficiency; they leave the excess DNA to neighbors or host cells to perform functions that they can take advantage of. There, also, is a tendency for increased cooperation to begin forming multi cellular creatures.
The much more complex individual eukaryote cell somehow carries within it the knowledge of differentiation into thousands of different cells each with intricate detailed tasks, such as neurons. This level of knowledge in one cell and the communication that develops between the individual cells in human beings is a staggering level of detailed cooperation.
But, somehow, even in much simpler bacteria, cooperation leads to versions of multi cellularity.