 Cells need to know where they are for many reasons. But, it is very challenging for an individual to know its exact location relative to a large outside world, without GPS. Despite great difficulties, remarkably, individual cells can make complex calculations and decisions based on their exact relations to other cells and organs.
Cells need to know where they are for many reasons. But, it is very challenging for an individual to know its exact location relative to a large outside world, without GPS. Despite great difficulties, remarkably, individual cells can make complex calculations and decisions based on their exact relations to other cells and organs.
Travelling white blood cells—T cells, B Cells, macrophages and leukocytes—need to know exactly where an infection is located and where they are in relation to it. Neurons in the fetus must find their exact places in enormously complex networks. Cells growing in an organ or limb must know exactly how they fit into the structure and shape and when to stop making new cells at the edge. How can these intelligent cells know their place exactly?
By following multiple rules and measurements and synthesizing this information, cells are able to calculate where they are. But, there are many possible sources of error that have to be taken into account.
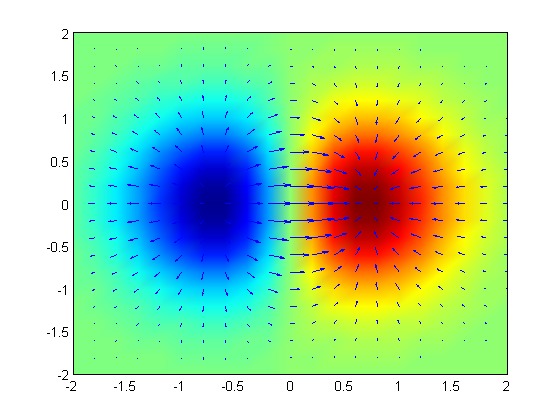
Cells use data from different sources such as morphogenic fields of diffusible molecules and electrical gradients and complex networks of genes. Recent research shows that as well as being able to calculate many different factors and taking them into account, cells are, at the same time, signaling back and forth to other cells, sharing information that helps determines the exact location.
While it might seem simple for cells to measure a gradient, in fact, it is not, because many variables can affect the measurement. To hone the accuracy of information from diffusing molecules, electrical gradients and mechanical signals, it must use feedback loops, multiple comparative measurements, timed measurements, complex gene regulation, principles of self-organization and very specific cell-to-cell signaling. Cells become veritable mathematicians to answer the questions. This post will describe some of the problems that cells must overcome to have accurate location information that is critical for brain structures and immune function.
Complex Travel and Cytokine Signals
 Immune cells that travel through the body to an inflammation site have to deal with a thousand different factors using many different mechanisms. Please see post the Complex Migration of Leukocytes for details. During this travel, cells need to know where they are and where they are going. A conclusion of that post was that cells rely upon very detailed signals, including cytokines, chemokines and neurotransmitters, and gradients from many different sources. Cells keep up chatter between other travelling immune cells, as well.
Immune cells that travel through the body to an inflammation site have to deal with a thousand different factors using many different mechanisms. Please see post the Complex Migration of Leukocytes for details. During this travel, cells need to know where they are and where they are going. A conclusion of that post was that cells rely upon very detailed signals, including cytokines, chemokines and neurotransmitters, and gradients from many different sources. Cells keep up chatter between other travelling immune cells, as well.
The tissues secrete attractant molecules that can help guide them through blood vessels, extracellular matrix and organs. For this type of behavior, immune cells have many different receptors that allow them to understand multiple signals. Each bodily tissue—lymph, liver, lung, skin and brain—also, has different scaffolds that must be traversed. The cells navigate using all of the gradients along with other mechanisms.
Many different gradients are produced that form waves of different types of information. Cells are able to analyze multiple competing signals and make decisions how to use them. Each signal triggers a cascade that stimulates specific genetic networks. But, some of the information is more accurate than others and decisions have to be made to determine the exact location.
Cell Signaling for Accuracy
New research is demonstrating the great complexity of cellular language. One type of communication between cells is touching and directly transporting data such as with synapses. Transmitted signals include immune cytokines and chemokines (attractants), neurotransmitters of many types, small and large RNAs, small peptides and hormones. Special plant signals are kinocidins. Signals can work either locally or at a very long distance. For the signal to work, the cells must manufacture the specific protein receptors and have them ready in the membranes.
Very recently, new types of communication have been discovered. It is not widely known that small nanotubes, built with actin molecules surrounded by membranes, are found between almost all cells. In fact, these cytonemes are the major way that viruses are transmitted between cells (see post). Cytonemes are used for horizontal gene transfer as well.
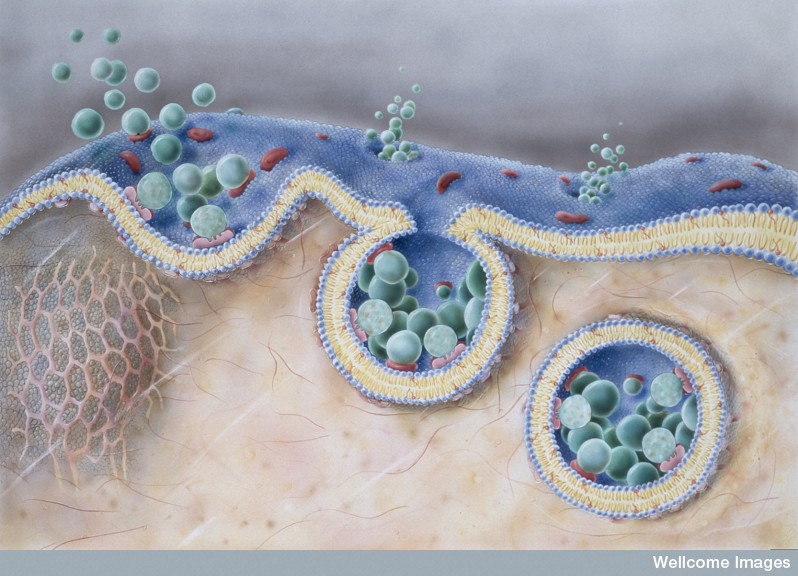 Another universal technique of cellular communication is vesicles. Exosomes are secreted from the membranes of cells and then taken in by other cells (as endosomes). It is now known that neurons and glia communicate with these vesicles and send important protein and genetic information including critical RNAs.
Another universal technique of cellular communication is vesicles. Exosomes are secreted from the membranes of cells and then taken in by other cells (as endosomes). It is now known that neurons and glia communicate with these vesicles and send important protein and genetic information including critical RNAs.
Even more unusual are the many different ways that cells use electricity and electric gradients. Please see the post on electrical gradients, electrical synapses and general electricity in the brain for details. An even more unusual technique is just being discovered for cellular communications using photons.
In each of these types of communication, the cell is looking for accuracy of measurement. They can compare these signals with different types of information to determine their accuracy.
Cellular Measurements of Location – The Need for Accuracy
 Measurements in the human world are made through measuring rods for short distances and radio waves for long distances. Cells use molecules that travel between cells for many local measurements. These molecules obey the simple laws of diffusion in a water solution. But, unlike radio waves, diffusion can occur at different speeds, moving slower as it spreads from the source. When the molecules form a stable gradient (a steady state), the cell can understand where they are in the field of the gradient. But, many gradients are not stable. So, the cell must devise diverse methods to ensure that their measurement is accurate.
Measurements in the human world are made through measuring rods for short distances and radio waves for long distances. Cells use molecules that travel between cells for many local measurements. These molecules obey the simple laws of diffusion in a water solution. But, unlike radio waves, diffusion can occur at different speeds, moving slower as it spreads from the source. When the molecules form a stable gradient (a steady state), the cell can understand where they are in the field of the gradient. But, many gradients are not stable. So, the cell must devise diverse methods to ensure that their measurement is accurate.
Many factors make gradient information unreliable. Molecules are manufactured at various rates and receptors are variable in activating signaling molecules and in their concentrations. The diffusible molecules are eliminated in different ways. They way a receptor operates can effect accuracy. Environmental factors such as temperature or rate limiting nutrients affect accuracy.
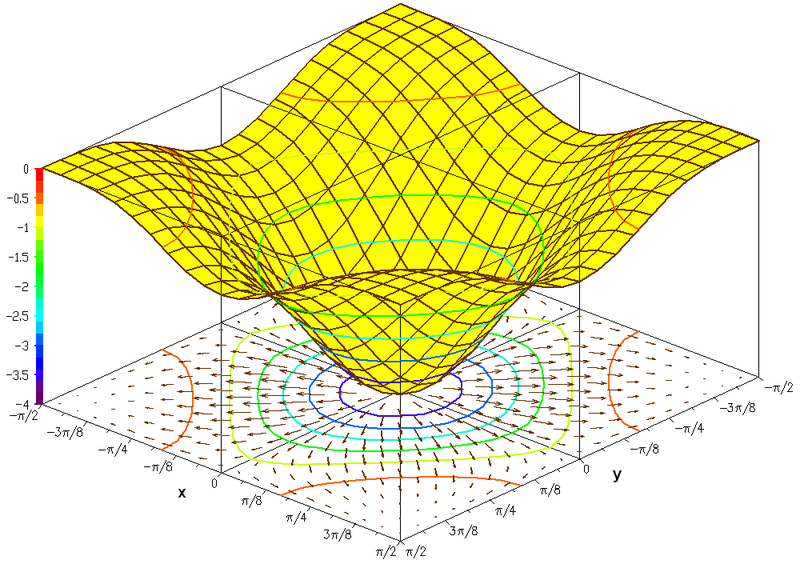 Other variables are the regulation of steps taken to produce and eliminate a molecule used for the gradient. The diffusible molecule can be made by the genes of the cell, or metabolized from other molecules in several steps.
Other variables are the regulation of steps taken to produce and eliminate a molecule used for the gradient. The diffusible molecule can be made by the genes of the cell, or metabolized from other molecules in several steps.
An example occurs in the embryo, where one gradient is retinoic acid. Retinoic acid forms the pattern upon which the back part of the brain is built. Retinoic acid is made in two metabolic steps from ingested vitamin A. This simple metabolic pathway is highly regulated in order to maintain the gradient. Maintaining the gradient involves feedback loops to the molecules in its synthesis, metabolic degradation and the receptor mechanisms in the cells. Special proteins are involved in regulating retinoic acid, as are the cytochrome P450 enzymes that eliminate it. When experiments alter the amount, these control pathways maintain the gradient and therefore the information the neurons use to build the brain tissue.
One set of errors occurs when environmental factors affect all cells in the same way, such as temperature. Other errors occur when each cell responds differently to an environmental factor.

Some cells can, by themselves, alter how the gradient is produced. A cell can, also, increase elimination of the gradient molecule. It can try to stabilize the signal by producing more receptors. But, these maneuvers produce other problems by having a saturation of receptors and further inaccuracy. The cell can take a measurement as the gradient develops, but this has to be timed exactly. Cells can, also, introduce another molecule that interacts with the diffusing signal.
Another way cells improve accuracy is to avoid measuring diffusion and, instead, use active transport. In plants there are no places between cells to diffuse and therefore active transport is a critical factor in determining where cells are. Water-soluble molecules are actively transported across plant membranes.
Two other situations create different measurement problems. Many small cells produce bigger fluctuations in measurement. A few big cells with a fixed larger numbers of receptors have problems of saturation.
In general, by improving any factor’s accuracy, others are made worse. To utilize gradients, specific factors must be emphasized that are the most reliable for comparison. Once the gradient is established at steady state, some reliable factors that don’t change diffusion include temperature, metabolic cell states, obstacles, and sticky objects. The cell can take these into account in different ways to firm up their measurements.
Quantity Measurements
 Cells can make multiple measurements and then synthesize these to use for positional information—either the same factor over time or multiple factors. Each of the calculations has to be independent of the others. In the posterior brain the fetal stem cells use not only retinoic acid for the gradient, but, also, use two other molecules at the same times—FGF and Wnt. Many other cells use multiple different diffusible gradients.
Cells can make multiple measurements and then synthesize these to use for positional information—either the same factor over time or multiple factors. Each of the calculations has to be independent of the others. In the posterior brain the fetal stem cells use not only retinoic acid for the gradient, but, also, use two other molecules at the same times—FGF and Wnt. Many other cells use multiple different diffusible gradients.
One technique to gather multiple measurements over time is to pick up the signal and place it in an endosome. Inside the cell in a vesicle, the signal can continue to be measured along with other signals.
 Another mechanism of counting is having a longer or shorter action at receptor complexes. When multiple signals interact with the complex before it responds, an average can be measured. Factors that disturb gradients for a short time will only affect some of the signals and the average would not be based on the short-lived disturbance.
Another mechanism of counting is having a longer or shorter action at receptor complexes. When multiple signals interact with the complex before it responds, an average can be measured. Factors that disturb gradients for a short time will only affect some of the signals and the average would not be based on the short-lived disturbance.
But, disturbances that last a long time need different mechanisms. This can occur when there are great differences in the number of cell receptors, whose manufacture takes a long time. In this situation, if the cell has to make the decision quickly, the information won’t work, but if it needs a longer time then the information can be accurate. Also, using timed averages implies that the gradient molecule has a short half-life. The cells have to wait a longer time to make their decision.
Some cells use a switch. Once switched on by a certain amount of signaling, it stays on until there is a major change to flip it off. This creates a time period that the signal is on.
During fetal development, stem cells often have to decide whether they will divide or not at a particular place. These types of decisions are made by a switch mechanism that has different thresholds. Many of these switches affect the change in genetic networks. These are dependent upon the amount of fluctuations that occur (noise).
Another technique is measuring the ratio of receptors that are triggered compared with those that aren’t (this is done with Hedgehog).
Using Other Cells for Measurement
 Cells that are near each other can add their information together. Individual cells make a measurement and then signal to neighboring cells about the impact on complexes inside the nucleus. One example occurs when forming a wing from a gradient signal. The signal triggers a transcription factor. At the edge of the forming wing there are very low signals that are inexact and don’t trigger the switch. Instead the cells communicate with each other in such as way that only cells are triggered when right next to a cell that is already triggered. This occurs, also, through timed switches. With these multiple signals, the wing is built out from the center to the edge, where it stops at an exact point.
Cells that are near each other can add their information together. Individual cells make a measurement and then signal to neighboring cells about the impact on complexes inside the nucleus. One example occurs when forming a wing from a gradient signal. The signal triggers a transcription factor. At the edge of the forming wing there are very low signals that are inexact and don’t trigger the switch. Instead the cells communicate with each other in such as way that only cells are triggered when right next to a cell that is already triggered. This occurs, also, through timed switches. With these multiple signals, the wing is built out from the center to the edge, where it stops at an exact point.
Other Problems and Solutions
 Cells are not always able to make independent measurements of multiple factors. All of the factors might be affected by one particular variable. If heat increases the density of the gradient, the cell can measure another molecule that is affected differently by the heat and compares them. These signals can then cancel each other out. This type of measurement occurs when certain factors have known affects that alter several gradient molecules being measured. If an environmental alteration can produce several different effects that are known, differences can be calculated to determine the location.
Cells are not always able to make independent measurements of multiple factors. All of the factors might be affected by one particular variable. If heat increases the density of the gradient, the cell can measure another molecule that is affected differently by the heat and compares them. These signals can then cancel each other out. This type of measurement occurs when certain factors have known affects that alter several gradient molecules being measured. If an environmental alteration can produce several different effects that are known, differences can be calculated to determine the location.
An example of using two signals occurs with the alteration of the rate of diffusion of the signal molecule and at the same time a change in the number of receptors in the cell that are picking up the signal. These two changes affect each other and the cell can make a complex determination from these measurements for the location. Somehow, cells are able to measure these mathematical abstractions. In the wing example given above the signal responds to the differences of cells to the gradient.
Some switches take account of multiple signals over a particular amount of time, but alter the answers based on how much random background noise there is. Understanding the amount of random signaling versus the real signal can be calculated.
Self Organizing Structures Among Multiple Cells
 Some signals only occur between local cells. But the signal can be powerful and spread from one local group to another as a moving front. Studies show that there is a limit to how far this type of information can spread, hence making a structure. Other rebounding gradients can then occur. The effect of these spreading waves can be measured as well and compared to a stable wave.
Some signals only occur between local cells. But the signal can be powerful and spread from one local group to another as a moving front. Studies show that there is a limit to how far this type of information can spread, hence making a structure. Other rebounding gradients can then occur. The effect of these spreading waves can be measured as well and compared to a stable wave.
An example of this spreading wave of information and signaling in human life occurs when we use the speed of light from lightning and compare it to the speed of thunder to determine how far away the lightning occurred. Cells make a similar calculation.
Multiple different interactions between local cells can create spatial ordering. Cells that are part of an initial signal, such as the Notch pathway, can participate in a second type of signaling as well. These secondary gradients organize more cells in different ways. One type of secondary signal can inhibit the original signal at an edge of the organ. Different levels of cell adhesion can be part of this signaling. Putting all of these together can create very organized patterns that are required in organs and the fetal brain.
Other location signals can be linked to these self-organizing systems as well. Self-organizing signals can, also, be correlated to a particular gradient signal noting the exact location on which to organize. The organization of the organ and the position sense of the individual cells can be part of the same larger process.
Intelligent Cells Know Their Place
 Cells need to know exactly where they are for many reasons—building the fetal brain, traveling to an infection site and maintaining bodily organs. They use many different types of information to answer this surprisingly complex question and must synthesize them all to make decisions. Problems arise with the accuracy of the information, such as using gradients made of diffusing molecules or electrical gradients. Each situation can involve many different variables that can be altered with temperature, metabolism and other environmental factors. Each cell must be able to use advanced mathematics for these determinations, extracting stable information from flux.
Cells need to know exactly where they are for many reasons—building the fetal brain, traveling to an infection site and maintaining bodily organs. They use many different types of information to answer this surprisingly complex question and must synthesize them all to make decisions. Problems arise with the accuracy of the information, such as using gradients made of diffusing molecules or electrical gradients. Each situation can involve many different variables that can be altered with temperature, metabolism and other environmental factors. Each cell must be able to use advanced mathematics for these determinations, extracting stable information from flux.
Other posts have described the complex language of cellular signaling using cytokines, chemokines, neurotransmitters, electrical signals, photon signals, nanotube signals, and vesicles filled with genetic information. Cells use all of these communication methods for comparing information with local cells. Nearby cells can start a new gradient to stop the process and mark an endpoint of the growth of an organ or limb.
How can an individual cell be so intelligent? How can an individual cell integrate so many different kinds of information and then act in concert with many other cells? Where does this intelligence lie? Where is the direction for all of this? The cell must be tied to other information sources such as mind.