 The common wisdom that myelin is just an insulator to make faster neuronal communication has been overthrown by recent research that finds much more complexity in how myelin is used by the brain. The old “neuron doctrine” (one of many useless dogmas) was that each neuron sends a signal to the next neuron, and so on, in computer like fashion. In fact, the process is vastly more complex and variable and the new myelin code adds to brain complexity in very important ways.
The common wisdom that myelin is just an insulator to make faster neuronal communication has been overthrown by recent research that finds much more complexity in how myelin is used by the brain. The old “neuron doctrine” (one of many useless dogmas) was that each neuron sends a signal to the next neuron, and so on, in computer like fashion. In fact, the process is vastly more complex and variable and the new myelin code adds to brain complexity in very important ways.
Research reported last week shows that myelin patterns are different in each brain region, especially each of the six layers of the cortex—myelin appears to have its own very complex code. Un-myelinated axons are crucial for newly discovered types of communication between the naked axons and adjacent local immune and nervous system cells. Both myelin, interspersed patterns of myelin, and lack of myelin are essential for specific types of information transfer. Previous posts have described the intelligent behavior of microglia and  astrocytes in creating, maintaining and pruning synapses. Now it appears oligodendrocytes, also, make many decisions concerning the types of signals.
astrocytes in creating, maintaining and pruning synapses. Now it appears oligodendrocytes, also, make many decisions concerning the types of signals.
Previous posts have noted that the neuron has many different methods of communication. In addition to the well-studied axon spikes triggering secretion of chemical neurotransmitters and the less studied electrical synapses and synchronous brain waves, there are now newly discovered sideways communication from the un-myelinated axons, nanotubes and vesicles. This can take the form of cytokine signaling to local immune cells and neurotransmitter signaling to other neurons, glia and immune cells. In addition, electrical information is transmitted in the form of local field potential with many contributing factors such as shape of cells, extracellular matrix and calcium spikes.
The new complex variations in the way myelin is used and not used will greatly complicate the “mapping of the brain,” because of the many newly discovered sideways types of signaling from the un myelinated axon.
Myelin Variations
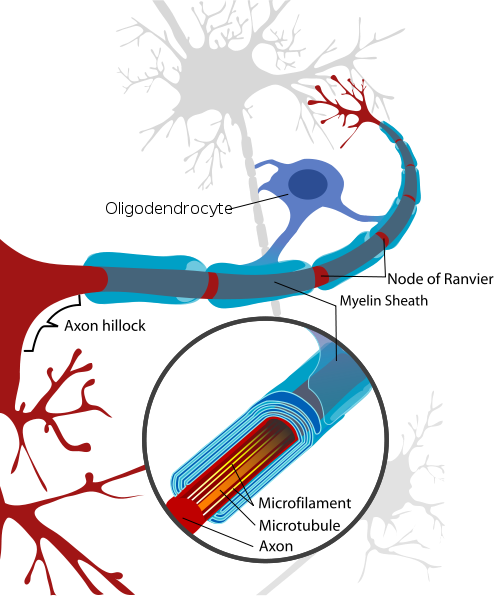 Until now, myelin has been considered to be just insulation for more rapid electrical current. But, in fact, axons are quite different from electrical wires. The new study shows that myelin affects the way information is created and communicated.
Until now, myelin has been considered to be just insulation for more rapid electrical current. But, in fact, axons are quite different from electrical wires. The new study shows that myelin affects the way information is created and communicated.
The study last week looked at myelin along axons in the six layers of the cortex. It found that in layers II and III, there are different patterns of very long intermittent stretches without any myelin. Also, the critical initial segment that determines the type and strength of the signal was different in each region. In fact, many different regions of the brain have different amounts and distribution of myelin. The myelin in each region is so different from each other, and so different than what was previously assumed, it appears to be yet another brain code.
Initial Segment Is Important and Varied
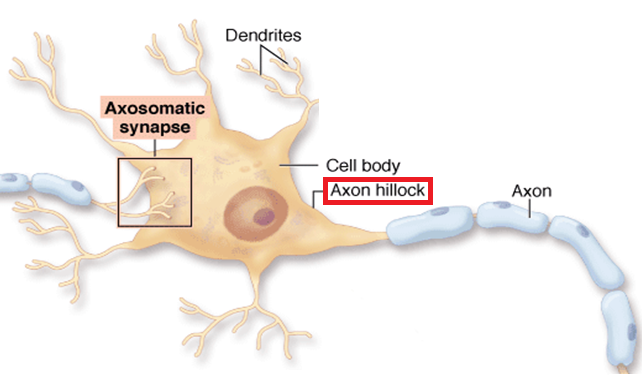 Perhaps, most significantly, the recent study found that the initial region from the cell body to the start of myelination along the axon was very different in each of the cortical layers. This initial segment is considered to be very important for the propagation of the signal and can vary from 5um to 80 um in length. It is where the decision is made whether or not to send an electrical signal along the axon. The initial segment has a very complex set of ion channels and structures that help make the decision to spike. It, also, determines the specific shape of the electrical spike and the amount of neurotransmitter release. The length of this segment was longer in layers III and IV than in V and VI.
Perhaps, most significantly, the recent study found that the initial region from the cell body to the start of myelination along the axon was very different in each of the cortical layers. This initial segment is considered to be very important for the propagation of the signal and can vary from 5um to 80 um in length. It is where the decision is made whether or not to send an electrical signal along the axon. The initial segment has a very complex set of ion channels and structures that help make the decision to spike. It, also, determines the specific shape of the electrical spike and the amount of neurotransmitter release. The length of this segment was longer in layers III and IV than in V and VI.
The initial segment, also, determines whether the spike can go backwards or not. Backward transmission determines aspects of neuroplasticity, such as in memory consolidation in the hipppocampus during deep sleep and during quiet reflection when awake.
Myelin, it now appears, also, plays a part in determining where synapses will occur. There are specific proteins in the myelin sheath that stop the budding of an axon. The oligodendrocyte and its myelin therefore determines the structure of the circuit.
Myelin in Higher Brain Centers
 Myelin is recent in evolution and was thought to, only, allow rapid communication of specific neurons for long communication. Myelination increases the speed eight times. But, many of the more recently evolved neurons, actually, have less myelin. The higher in the cortex, the less myelin there is. Also, the neurons of the cortex and higher regions use myelin in totally different ways. This is a form of “intermittent myelin” where there are myelin regions and then long tracts on the same axon with no myelin in new and complex patterns.
Myelin is recent in evolution and was thought to, only, allow rapid communication of specific neurons for long communication. Myelination increases the speed eight times. But, many of the more recently evolved neurons, actually, have less myelin. The higher in the cortex, the less myelin there is. Also, the neurons of the cortex and higher regions use myelin in totally different ways. This is a form of “intermittent myelin” where there are myelin regions and then long tracts on the same axon with no myelin in new and complex patterns.
The classic view has been a consistent pattern of long tracts of myelin with small breaks (nodes) to allow rapid electrical jumping from node to node. The assumption has always been that this occurred everywhere, but it turns out it doesn’t.
The higher-level neurons that are considered to calculate the most advanced material use myelin sparingly and in different amounts. These large regions without myelin allow communication sideways from the neurons to neighboring neurons and other cells.
There appears to be a dialogue between the oligodendrocyte and neuron concerning the appropriate amount of myelin to be used. This is not really surprising when we consider the constant signaling and communication between the other two glial cells and the neuron—see posts on astrocytes determining synapses and the microglia patrolling and pruning neurons and synapses.
It has been recently shown that neuronal activity stimulates stem cells to make more oligodendrotyes, which increases myelination in the premotor cortex and the subcortical white matter. These signals improved motor functions in correlated limbs.
Myelin
 Myelin is a characteristic of vertebrates, but through parallel evolution some invertebrates, also, use a different type of myelin. Myelin begins in the 14th week of the fetus and few neurons are myelinated at birth. Myelination continues gradually providing a baby’s physical capacities—3 months moving head, 6 months learning to sit, and 7 to 12 months crawling to walking. Myelination continues to increase throughout childhood and in late adolescence and early adulthood the pre frontal cortex myelinates, correlated with increased planning and executive ability and less impulsiveness.
Myelin is a characteristic of vertebrates, but through parallel evolution some invertebrates, also, use a different type of myelin. Myelin begins in the 14th week of the fetus and few neurons are myelinated at birth. Myelination continues gradually providing a baby’s physical capacities—3 months moving head, 6 months learning to sit, and 7 to 12 months crawling to walking. Myelination continues to increase throughout childhood and in late adolescence and early adulthood the pre frontal cortex myelinates, correlated with increased planning and executive ability and less impulsiveness.
In the periphery Schwann cells produce the myelin and in the brain oligodendrocytes. There are different cell subtypes that make slightly different myelin in various regions. Myelin includes fat for insulation and therefore looks like white sausage—the white matter of the brain. Myelin consists of 40% water and the portion that is not water is 80% fat (mainly galacterocerebroside including cholesterol) and 20% proteins (myelin basic protein, oligodendrocyte protein, and proteolipid protein). Strengthening molecules include sphingomyelin hydrocarbon chains.
Myelinated Neurons
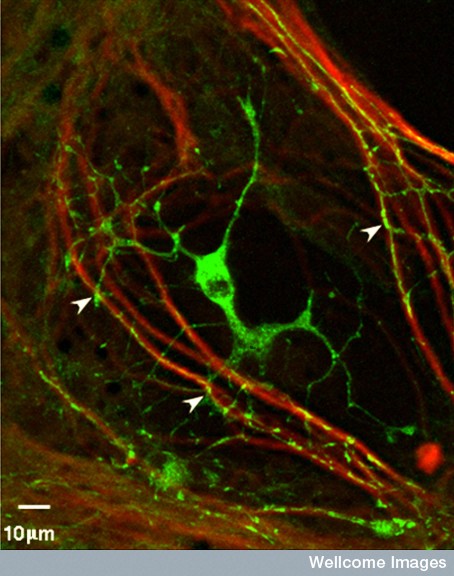
Myelin consists of cell membranes compacted and wrapped multiple times around the axon. This membrane is formed by oligodendrocytes. The oligodendrocyte first sends small arms to touch and grab the axon (see picture on right of a green oligodendrocyte grabbing an axon). Then they wrap the axon with 20 to thirty layers of membrane. Many oligodendrocytes grab one axon along its entire length and leave spaces between each arm. These areas of exposed axon have a larger amount of sodium channels that respond to voltage change of 20mV by rapidly opening for sodium to enter the cell causing the action potential. With the myelin in the traditional pattern, the action potential only occurs at these nodes (of Ranvier) and then jumps to the next node.
In un-myelinated axons, action potentials travel slowly down the entire axon. Factors that affect the speed are the amount of myelin wrapping, the size of the un-myelinated nodes, and the length of the myelin along the axon.
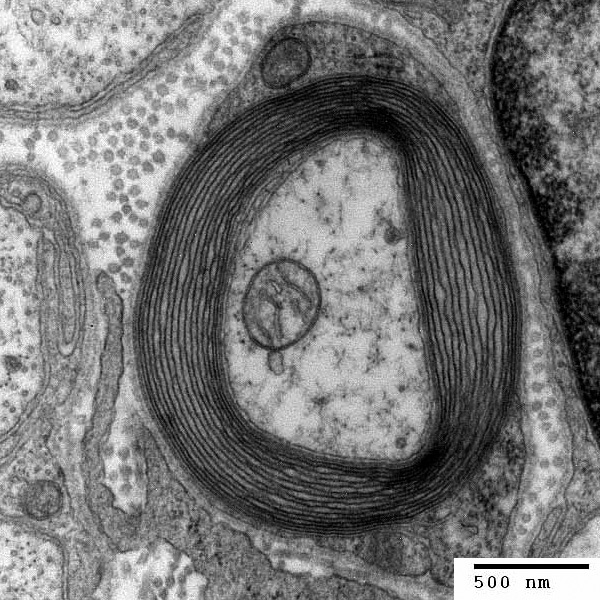 The differences in the amount of myelin on the axon discovered this week could be related to various different speeds necessary for different parts of the cortex. For example, it could vary based on the time of arrival to some part of a circuit. Very long nodes can greatly alter and slow down the transmission. The structures of the channels in the un-myelinated sections are not known but could also be critical in determining the function.
The differences in the amount of myelin on the axon discovered this week could be related to various different speeds necessary for different parts of the cortex. For example, it could vary based on the time of arrival to some part of a circuit. Very long nodes can greatly alter and slow down the transmission. The structures of the channels in the un-myelinated sections are not known but could also be critical in determining the function.
These differences could be determined by the neuron or by local electrical, chemical and extra cellular matrix properties of the region. It could also be determined by the oligodendrocyte, which produces smaller nodes later in life. The cortical layering structure appears to be critical in these differences, because when the layering is disrupted, so is the pattern of myelination differences between the layers.
The important finding in this recent study is that there is yet another code—the myelin code. (Previous posts have noted the multiple genetic codes, as well as the different types of neurons and synapse. See post on more Genetic Complexity).
New Mechanisms of Myelin Stimulation
Recently, a variety of different ways that myelin is stimulated have been discovered.
1. One is dependent on glutamate stimulating NMDA receptors on stem cells that make oligodendrocytes. Neuregulin appears to switch to this mode by increasing these receptors.
It has been noted, also, that glutamate is increased in sleep (especially REM). Also, in sleep stem cells are stimulated to make oligodendrocytes and, therefore, more myelin. An opposite effect occurs while awake — less apoptosis and less oligodendrocytes. This is yet another great benefit of sleep.
2. Previous posts showed that microglia survey specific territory of the brain tapping on neurons in that region. Now it has been shown that precursor cells (NG1 cells) that make oligodendrocyte, also, continuously travel in a local environment with moving filopodia and large growth cones ready to make more cells. The exact density of these stem cells is maintained through a balance of active growth, self-repulsion by other stem cells, rapid movement and cell death. When damage occurs, these cells rapidly move to the injury to help repair neurons.
3. Recently, the well-known anticholinergic medication benztropine (used for Parkinson’s) was shown to promote stem cells to make oligodendrocytes and myelin. The exact mechanism is not yet clear.
4. When oligodendrocytes are old they tend to make less myelin. It was recently shown that this not from being old, but from lack of stimulating factors from other regions of the body.
5. Yet another mechanism is from specific types of M2 microglia and immune macrophages stimulating increased myelin.
Importance of Unmyelinated Neurons and Segments
There are a large number of axons that do not have myelin. In fact, there are advantages of neuron’s not being myelinated.
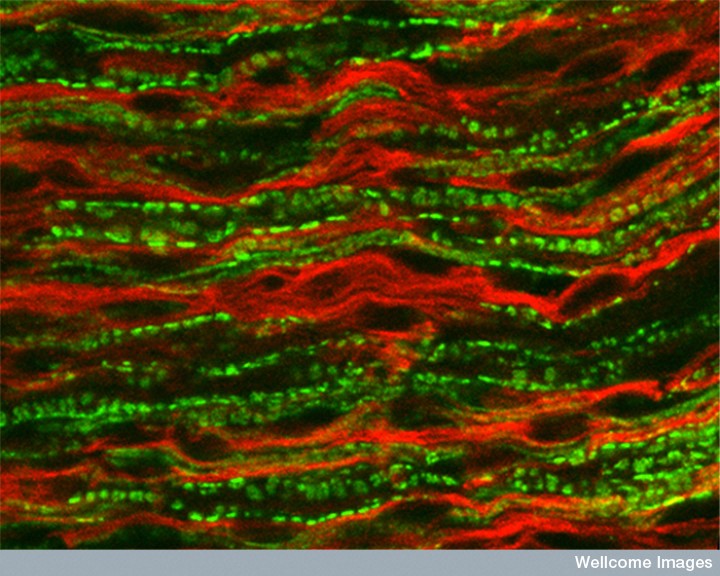
The newly described local lateral communication from axons can only occur in completely unmyelinated neurons or sections that are un-myelinated. These include the vital cytokine communication with immune cells in inflammation.
Axons travelling to and from internal organs in sympathetic, parasympathetic and hormone networks are largely unmyelinated, or only slightly myelinated. “Slightly myelinated” means that only one Schwann cell attaches to many different neurons. These circuits are vital for the normal homeostasis of all internal organs and appears to need these newly discovered sideways methods of communication.
 The internal milieu greatly benefits from un-myelinated axons’ sensing chemical changes in the environment, as well as local immune cytokine signaling. Without myelin the entire axon can respond to the extracellular matrix, energy flow and ionic status (Please see a previous post Brain Electricity and the Mind.)
The internal milieu greatly benefits from un-myelinated axons’ sensing chemical changes in the environment, as well as local immune cytokine signaling. Without myelin the entire axon can respond to the extracellular matrix, energy flow and ionic status (Please see a previous post Brain Electricity and the Mind.)
Previous posts on the relation of the immune and nervous systems showed that the vagus nerve is critical in connecting nervous system circuits with vital immune functions. These functions occur all along the un-myelinated vagus nerve. There is now a large amount of data showing immune communicating with back and forth signals to the vagus nerve about local immune activity. This local communication is, actually, vital for regulation of inflammation. There are a large number of special receptors and neurotransmitter signals along the vagus for this sideways local communication, including receptors for ATP, serotonin, acetylcholine and capsaicin. None of this could happen with myelin.
Un-myelinated neurons are now know to, also, communicate with cytokine signals to immune cells, sideways secretion of neurotransmitters to adjacent neurons and immune cells, and with vesicles filled with genetic and protein information and nanotubes. The study of these subtle recently discovered forms of communication is just beginning.
New Myelin Code Adds To Brain Complexity
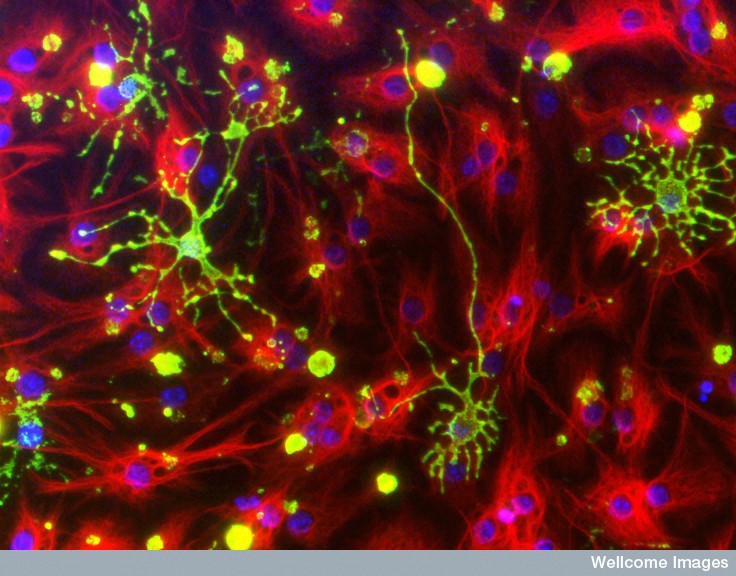 Previous posts show the critical parts that astrocytes and microglia play in the production, maintenance and pruning of the neuronal network. Now oligodendrocytes are shown to be a much more active player, as well.
Previous posts show the critical parts that astrocytes and microglia play in the production, maintenance and pruning of the neuronal network. Now oligodendrocytes are shown to be a much more active player, as well.
Myelin deposition is not a passive simple process. The structures of myelin, the number of layers of myelin, the length of the axon’s initial segment, the different lengths of the myelin in each part of the neuron, the size of the nodes, and the large areas of unmyelinated axons, are all critical to determine the function of the neuronal network. Myelin is not a simple insulator. It organizes the neuronal network and the types of information transfer. It allows areas for critical sideways local communication between neurons and immune cells. It determines the speed and flow of circuits.In fact, myelin forms a new critical code in the brain.
When looking at the many overlapping codes in the brain — complex patterns of gene networks, alternate RNA splicing, different neuron types, different synapse types, different types of information transmission, unique patterns of myelin — it is reasonable to ask where the direction for all of this is coming from?