 How is thought translated into molecular changes in brain cells? With constant changes to the wiring of the brain from mental activity, it is striking how many different ways the molecules in the circuits and synapses respond in milliseconds to new learning and experiences. Each day new neurons are minted in the hippocampus and integrated into the learning circuits. Triggers for increased new cells include learning, focused concentration, exercise, enriched environments, medications, psychotherapy, and meditation. Minute-by-minute new circuits are built, strengthened and altered. Each night unnecessary circuits are pruned. There are many different molecular mechanisms to build and strengthen synapse and wide circuits. Now, there is another form of neuroplasticity by switching glutamate NMDA subunits. This may be the most significant of all recently discovered mechanisms because of the importance of this type of receptor in the entire brain.
How is thought translated into molecular changes in brain cells? With constant changes to the wiring of the brain from mental activity, it is striking how many different ways the molecules in the circuits and synapses respond in milliseconds to new learning and experiences. Each day new neurons are minted in the hippocampus and integrated into the learning circuits. Triggers for increased new cells include learning, focused concentration, exercise, enriched environments, medications, psychotherapy, and meditation. Minute-by-minute new circuits are built, strengthened and altered. Each night unnecessary circuits are pruned. There are many different molecular mechanisms to build and strengthen synapse and wide circuits. Now, there is another form of neuroplasticity by switching glutamate NMDA subunits. This may be the most significant of all recently discovered mechanisms because of the importance of this type of receptor in the entire brain.
 Most mental events trigger signals in wide areas of the brain in milliseconds, and neuroplasticity occurs in these distributed circuits throughout the brain. Importantly, many different molecular mechanisms operate at the same time throughout these circuits. Previous posts have discussed how the more widespread and complex the neuronal circuit involved in the experience or learning, the more powerful it is. See previous post Neuroplasticity Primer and Update for details.
Most mental events trigger signals in wide areas of the brain in milliseconds, and neuroplasticity occurs in these distributed circuits throughout the brain. Importantly, many different molecular mechanisms operate at the same time throughout these circuits. Previous posts have discussed how the more widespread and complex the neuronal circuit involved in the experience or learning, the more powerful it is. See previous post Neuroplasticity Primer and Update for details.
A New Glutamate Form of Neuroplasticity
 Glutamate, while not as widely recognized in the popular press as many other better-known neurotransmitters, is actually, by far, the predominant neurotransmitter in the brain. One important form of neuroplasticity has been known for many years involving the glutamate receptor AMPA. In early research on long-term potentiation (LTP), stronger synapses are correlated with an increase of the number of AMPA receptors.
Glutamate, while not as widely recognized in the popular press as many other better-known neurotransmitters, is actually, by far, the predominant neurotransmitter in the brain. One important form of neuroplasticity has been known for many years involving the glutamate receptor AMPA. In early research on long-term potentiation (LTP), stronger synapses are correlated with an increase of the number of AMPA receptors.
Now, a totally new form of neuroplasticity has been discovered in the NMDA glutamate receptor that is closely connected with the AMPA receptor.
Previously Described Types of Neuroplasticity
There is a staggering amount of different ways that the neuronal circuits instantly change in response to learning. The following is a partial simplified list:
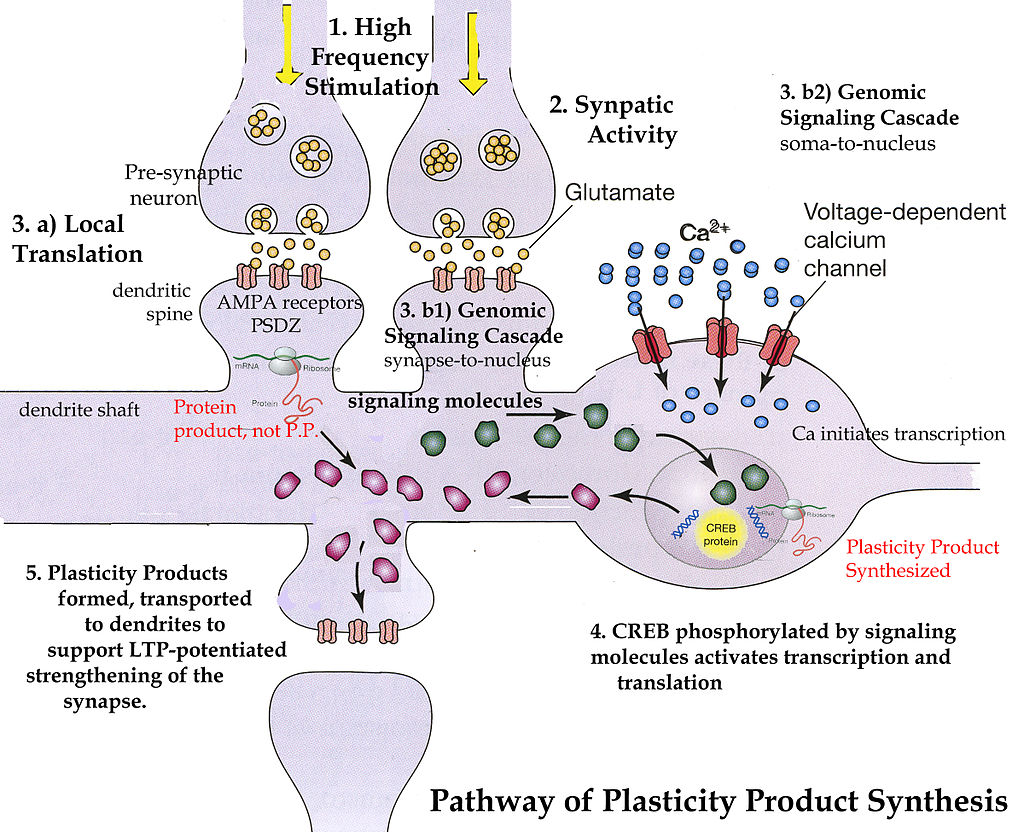 1. The type of neuroplasticity most studied as a cause of LTP involves a complex, and unusual, synapse where the pre synaptic neurons release both glutamate and another neurotransmitter, often glycine, and must trigger two different postsynaptic receptors, the AMPA and the NMDA. When both the AMPA and NMDA receptors are triggered, the NMDA receptor opens and calcium rushes in. This calcium influx triggers the manufacture of more AMPA receptors, which are placed in the synapse, and this results in an increase in the size of the signals (LTP). More calcium, more AMPA receptors cause a lasting increase in the signal called long term potentiation. This type of neuroplasticity has been most closely associated with declarative conscious memory and learning.
1. The type of neuroplasticity most studied as a cause of LTP involves a complex, and unusual, synapse where the pre synaptic neurons release both glutamate and another neurotransmitter, often glycine, and must trigger two different postsynaptic receptors, the AMPA and the NMDA. When both the AMPA and NMDA receptors are triggered, the NMDA receptor opens and calcium rushes in. This calcium influx triggers the manufacture of more AMPA receptors, which are placed in the synapse, and this results in an increase in the size of the signals (LTP). More calcium, more AMPA receptors cause a lasting increase in the signal called long term potentiation. This type of neuroplasticity has been most closely associated with declarative conscious memory and learning.
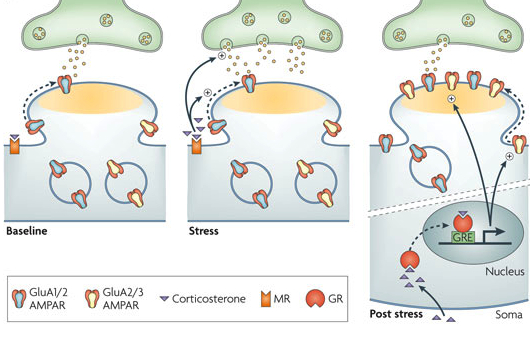
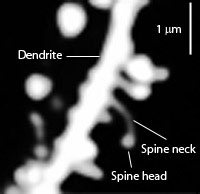 2. Another form of neuroplasticity is where the dendrite changes structure, specifically the size of the head of the dendrite spine. This increases the ability to rapidly receive information and makes it more stable. Spine alterations are influenced by, at least, BDNF and estrogen. It occurs as part of neuroplasticity in multiple parts of large circuits, for example, those that include the hippocampus and amygdala at the same time. Stress stimulates this type of neuroplasticity reducing the size of the hippocampus and increasing the amygdala. These alterations are influenced by environment, seasonal changes, and age.
2. Another form of neuroplasticity is where the dendrite changes structure, specifically the size of the head of the dendrite spine. This increases the ability to rapidly receive information and makes it more stable. Spine alterations are influenced by, at least, BDNF and estrogen. It occurs as part of neuroplasticity in multiple parts of large circuits, for example, those that include the hippocampus and amygdala at the same time. Stress stimulates this type of neuroplasticity reducing the size of the hippocampus and increasing the amygdala. These alterations are influenced by environment, seasonal changes, and age.
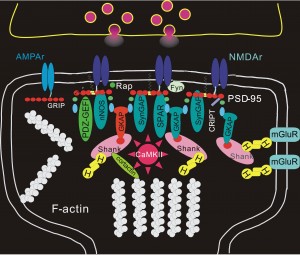 3. Complex alterations in the postsynaptic density, PSD (a structure of more than 1000 interlocking proteins which has different composition in different types of neurons), also are related to LTP.
3. Complex alterations in the postsynaptic density, PSD (a structure of more than 1000 interlocking proteins which has different composition in different types of neurons), also are related to LTP.
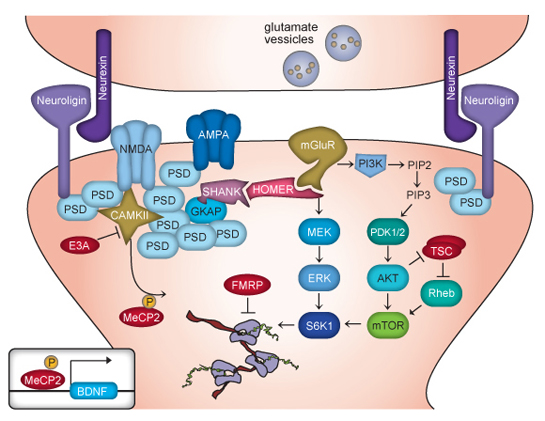
4. Another form alters large molecules that stick out from either side of the synapse into the space between the pre and post-synaptic cells. Significantly, neuroligins on the post-synaptic membrane meet the b-neurexins from the presynaptic membrane and shake hands, holding the synapse together.
5. A surge in calcium in neurons can trigger new “memory proteins”.
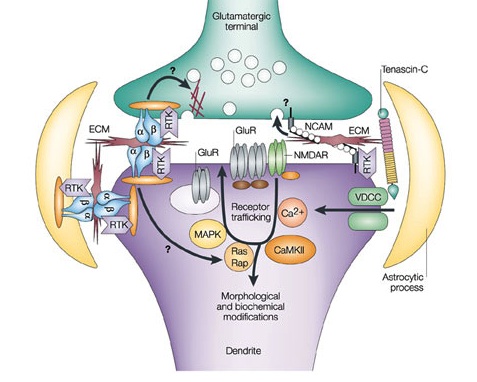 6. An unusual form was recently discovered where presynaptic neurons change their neurotransmitters, and the postsynaptic neuron changes its receptors to match the new neurotransmitter. This was discussed in a previous post and is based upon the fact that all neurons have the genetic machinery to make all of the neurotransmitters.
6. An unusual form was recently discovered where presynaptic neurons change their neurotransmitters, and the postsynaptic neuron changes its receptors to match the new neurotransmitter. This was discussed in a previous post and is based upon the fact that all neurons have the genetic machinery to make all of the neurotransmitters.
7. Alterations in signaling pathways from the membrane to the nucleus occur in various forms of neuroplasticity.
9. Transcription factors and RNA’s noted in a previous post, are also critical in some neuroplasticity.
10. The balance of inhibition and stimulation from many important brain circuits including the neurotransmitters, acetylcholine, serotonin, dopamine can also change the plasticity.
11. Intrinsic neuroplasticity involves an alteration of the membrane and the ion channels along the neuron that create the electrical signal. Alterations of sodium and potassium channels change the electrical wave along the axon. This type occurs especially in cerebellar motor learning.
12. Many different types of inter-neurons in the cerebellum (Golgi, unipolar, Lugaro) all have multiple mechanisms of neuroplasticity, which change the synapses.
13. Some circuits have multiple changes in different parts of the synapse at the same time (electrical, ionic channels, signaling molecules)
14. A totally different complex set of mechanisms are involved in the climbing fibers of the cerebellum, the major fibers that connect vision, balance and movement.
Glutamate Receptors Are Used in up to 80% of All Synapses
 Glutamate neurons are responsible for most of the brain function related to learning and memory. Glutamate is used by up to 50% to 80% of all neurons and 80% of all synapses in the brain use glutamate (many synapses use multiple neurotransmitters). Also, 80% of all the energy used by the brain is consumed in glutamate activity.
Glutamate neurons are responsible for most of the brain function related to learning and memory. Glutamate is used by up to 50% to 80% of all neurons and 80% of all synapses in the brain use glutamate (many synapses use multiple neurotransmitters). Also, 80% of all the energy used by the brain is consumed in glutamate activity.
Glutamate neurons are the backbone of the brain with by far the greatest amount of excitatory neurons. Another 35% of the brain are inhibitory neurons (GABA and glycine), which counterbalance the excitatory neurons. Interestingly, GABA is made in chemical reactions from glutamate. This counter balance of excitatory and inhibitory is critical since too much glutamate is toxic and causes destruction of cells (which is exactly what happens in a stroke when too much glutamate is released).
While most people have heard of dopamine, serotonin, acetylcholine, norepinephrine and histamine neurons, these all comprise a tiny percentage of the neurons and are basically modulating the brain structure of glutamate and GABA activity. The reason that there has been little media about glutamate is that it is much more complex and therefore there is much more to learn. In fact, much of the psychiatric theory based on the other neurotransmitters is probably incorrect; the mechanisms are really glutamate with some modulation by the other neurotransmitters. An entire new generation of medications awaits the elucidation of the monumentally complex glutamate and GABA structure. But, there are so many different types of glutamate receptors in different parts of the brain the research has been extremely difficult.
But, one thing is certain. All cognitive ability is dependent upon glutamate neurons, and especially NMDA.
Basic Glutamate and NMDA
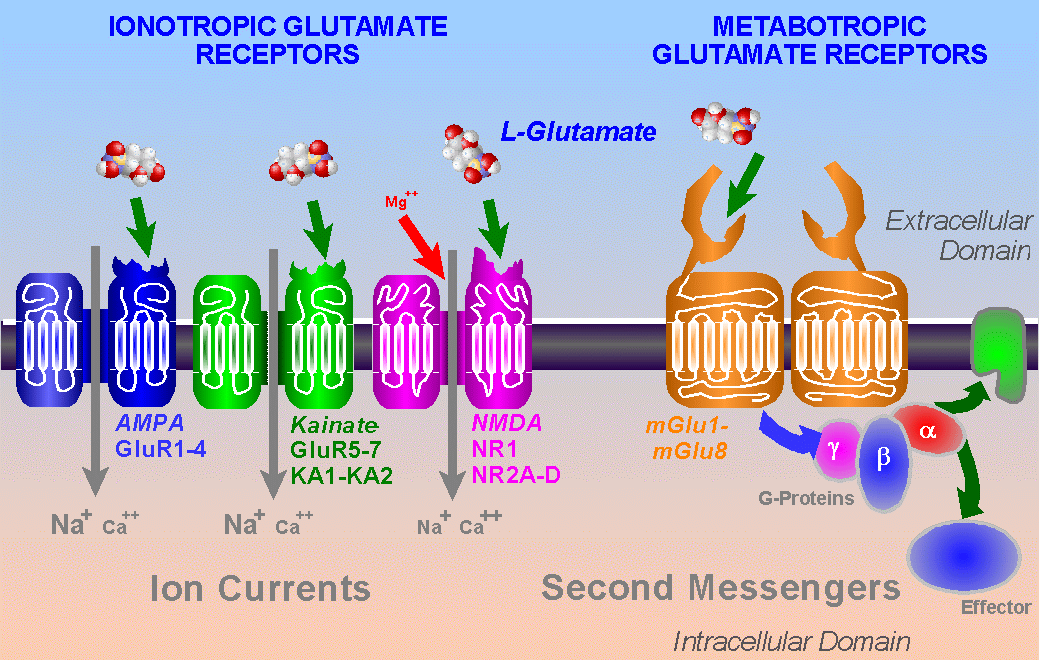 There are two basic types of receptors in the brain; one uses channels for chemical ions that open and close (inotropic) and the other are large proteins that create a cascade of signaling molecules from the membrane down to the nucleus (metabotropic). Different glutamate receptors use both mechanisms. Here we discuss only the ion channel type of which there are three:
There are two basic types of receptors in the brain; one uses channels for chemical ions that open and close (inotropic) and the other are large proteins that create a cascade of signaling molecules from the membrane down to the nucleus (metabotropic). Different glutamate receptors use both mechanisms. Here we discuss only the ion channel type of which there are three:
- AMPA
- Kainate
- NMDA
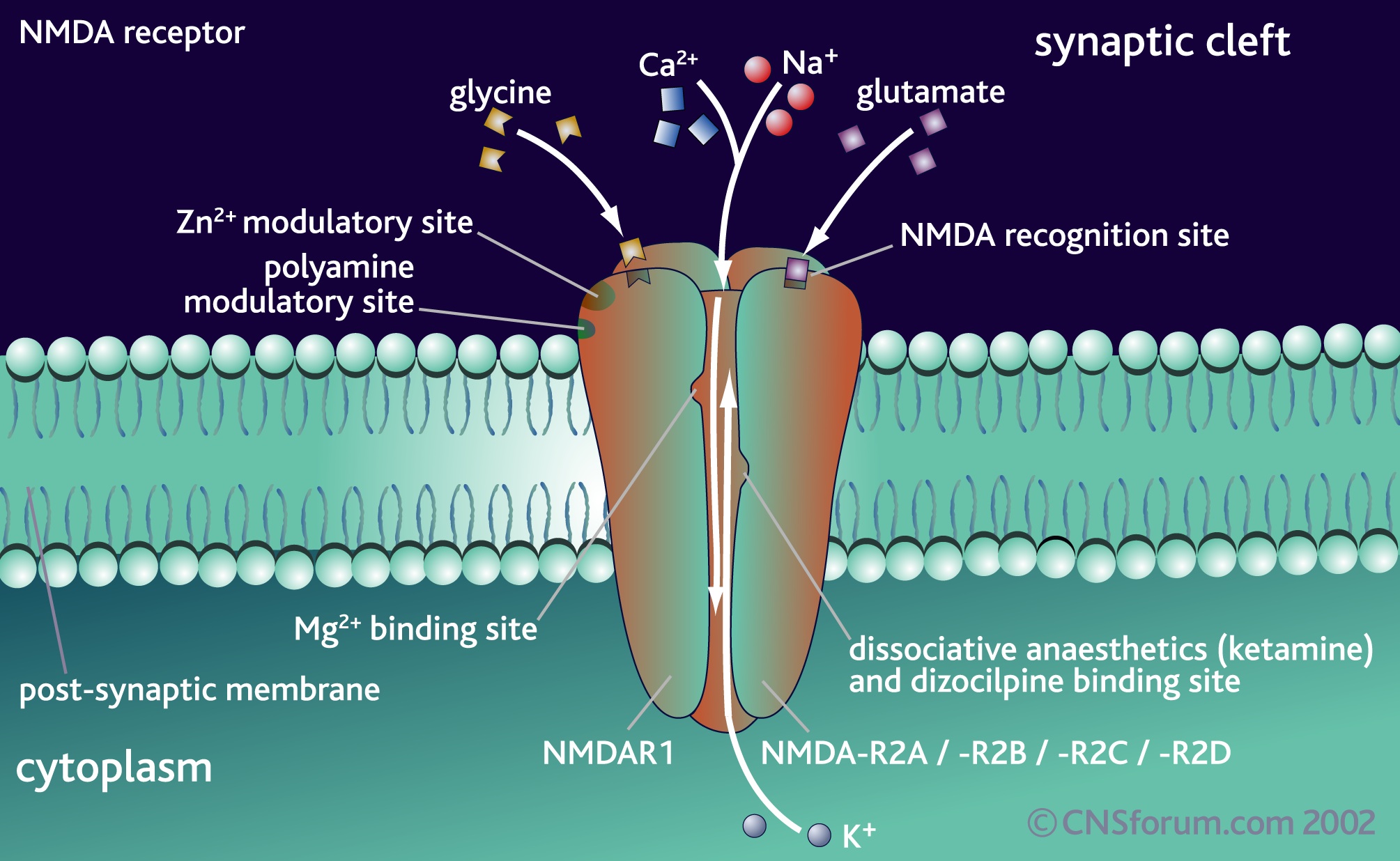 Of these, NMDA are extremely significant for all learning. It was mentioned above that in order to trigger the NMDA there needs to be two different neurotransmitters.
Of these, NMDA are extremely significant for all learning. It was mentioned above that in order to trigger the NMDA there needs to be two different neurotransmitters.
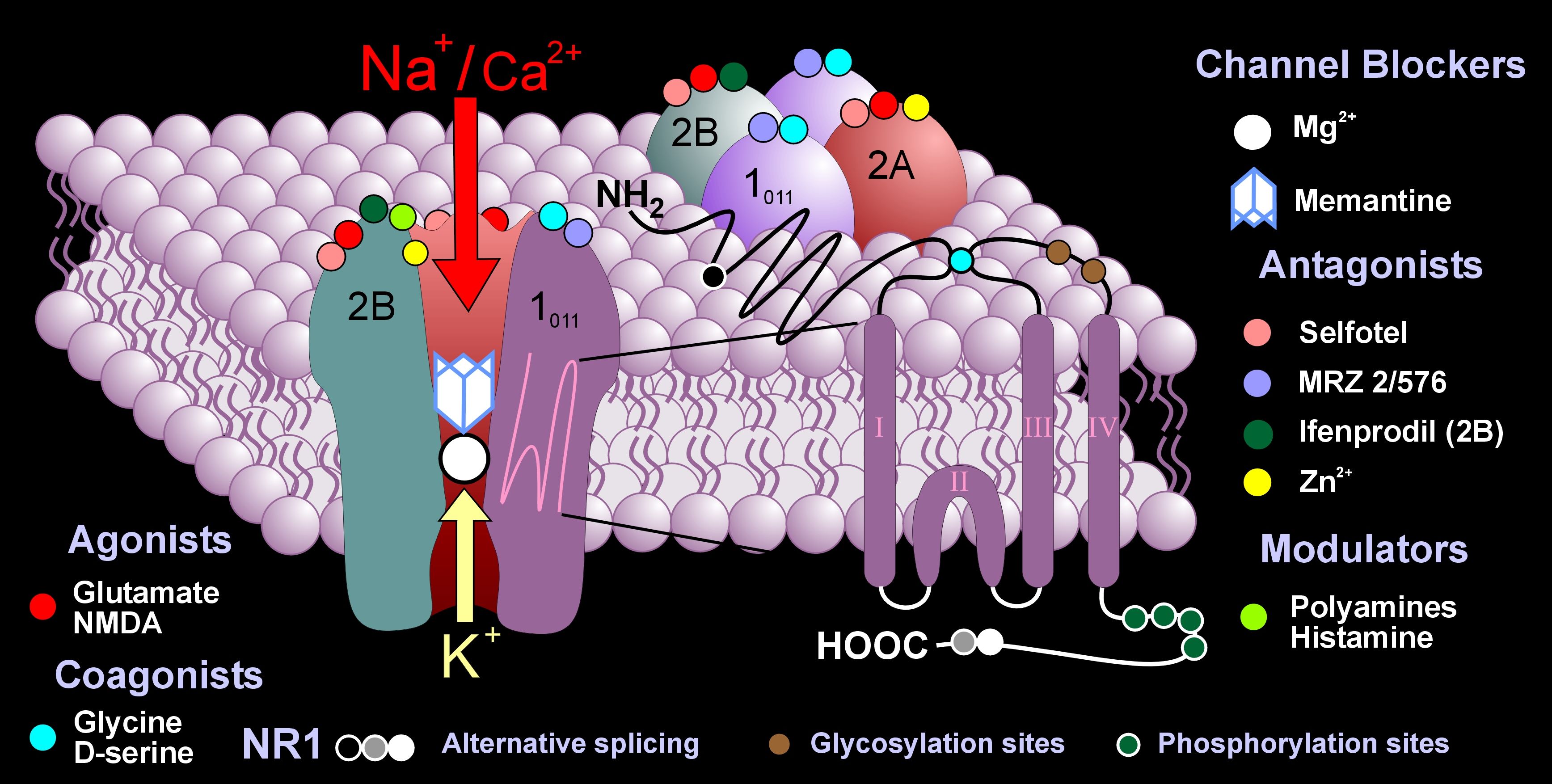
NMDA is, also, unique in that it has many other places on the receptor for other chemicals to interact with it and alter or modulate it in various ways.
NMDA has many other unique qualities.
 It is normally blocked by a magnesium atom that has to be released to open the channel. Plasticity is determined by how much calcium can get through after moving the magnesium.
It is normally blocked by a magnesium atom that has to be released to open the channel. Plasticity is determined by how much calcium can get through after moving the magnesium.
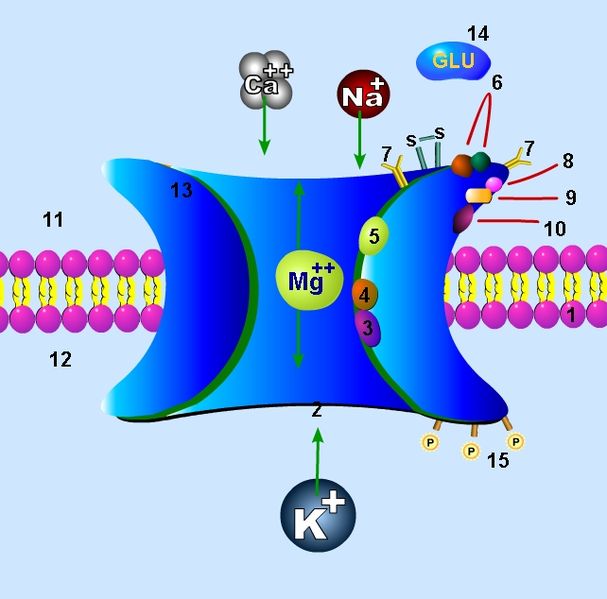
- Also uniquely, NMDA is influenced by voltage (the amount of ions on the inside and outside) but also by a neurotransmitter. All other receptors function with one or the other mechanism.
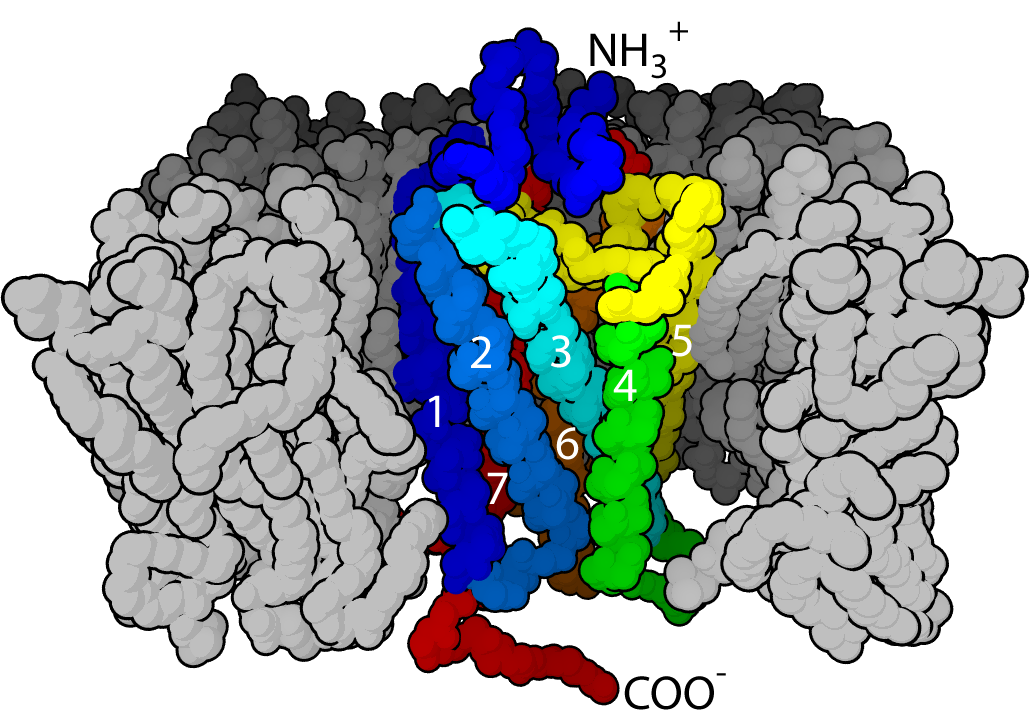
Another unique characteristic of NMA is that it is built from a variety of subunits, which are different in different parts of the brain. In fact, there are at least seven different subunits spread out through the adult brain. But, there may actually be many more since it is impossible to find all of the subunits, which are often used in small areas. This large number of different types each has distinctive properties in signaling, pharmacology and physiology.
Pharmacologically, NMDA receptors are very responsive to a large number of molecules because of the large number of extra binding sites, which are different with each subunit. Many chemicals that are normally in the brain effect these modulation sites such as polyamines, protons, and Zn, all strongly influenced by subunits.
But, the most shocking finding is that the subunits can switch in and out of the receptor changing its characteristics. And changes in a NMDA subunit are the newly discovered form of neuroplasticity. This could be the most significant type of neuroplasticity since there are so many places in the brain that use NMDA.
The different subunits appear to alter the amount and the timing of the calcium flow, which is related to the plasticity. But, now it appears that a critical factor is the specific subunits in the receptor that give it special calcium transfer abilities and specific signaling cascades.
NMDA Subunits
There are three families of subunits.
- One N1 subunit
- Four different N2 subunits (N2A, B, C and D) all made by completely different genes
- Two N3s (N3A and N3B) also from different genes.
 Each NMDA receptor is built from four subunits that are able to switch.
Each NMDA receptor is built from four subunits that are able to switch.
Usually it comprises two N1 and either two N2, or a combination of N2 and N3. In the fetus there are only N2B and N2D then others are added in the first two weeks of life. After a while N2A becomes dominant in the entire nervous system. N2D ends up being only in the diencephalon and mesencephalon, and N2B only in the forebrain, N2C only in the cerebellum and olfactory bulb. In the adult, N2A and N2B are predominant in the hippocampus and cortex, and important for plasticity and creation of synapses.
The Language of NMDA Subunits
Double receptors use only two types of subunits, triple use three.
The varieties and characteristics of subunits are just being unearthed.
- A triple structure with N1/N2A/N2B is very prominent in the hippocampus and cortex and possibly are 15% to 50% of all glutamate receptors. There are other double and triple combinations as well.
- There are many very specific differences such as ganglion cells that are “On” use N2B and “Off” N2A. These differences affect both the timing of synapses and the way signals are summed.
- N2A is very important for LTP at CA3-CA1 synapses in the hippocampus. And N2B seems to confer the opposite LTD in some cases, but not always. Other regions use different combinations.
- During sensory experiences the amount of N2A and N2B shift in minutes. Metaplasticity is also related to these subunits. (see post describing metaplasticity).
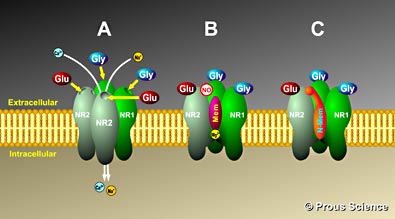
There are also big differences in the composition of receptor subunits based upon where the receptor sits on the neuron or astrocyte. Those, for example, in post synaptic sites are double N1/N2A and triple N1/N2A/N2B. Those receptors that are not near the synapse have N2B. But, in fact some neurons have the opposite, or other combinations.
The Subunits are Mobile
 These subunits of the receptor appear to be mobile and can exchange from the receptors in the synapse and those nearby. As well as NMDA being the predominant post synaptic receptors they can also exist on the pre synaptic neurons modulating the synapse.
These subunits of the receptor appear to be mobile and can exchange from the receptors in the synapse and those nearby. As well as NMDA being the predominant post synaptic receptors they can also exist on the pre synaptic neurons modulating the synapse.
NMDA receptors are also frequent on glial cells, the astrocytes, which control all aspects of the neuronal synapses (see post) and the oligodendrocytes, which make myelin. These are often different from the neuronal combinations including N2C and N2.
NMDA Subunit Structure and Properties
Subunits are divided into four regions: related to assembly of the receptor; binding glutamate and the second neurotransmitter glycine; regulating the pore and ion selectivity; and sending information to the cascade of molecules down to the nucleus and the DNA.
 Perhaps the most remarkable difference from other receptors is the tremendous variation in physical, pharmacological and signaling characteristics determined by the different subunits.
Perhaps the most remarkable difference from other receptors is the tremendous variation in physical, pharmacological and signaling characteristics determined by the different subunits.
Subunits dramatically change the properties of the magnesium and calcium effects on the receptor. In the different regions of the brain, signaling can be from high or low frequency determined by the subunits. Another type of subunit in the myelin making cells allows the receptors to be active even though there might not be much spiking in the myelin.
The subunit will determine how large the channel opens, and how sensitive the receptor is to the transmitters, glutamate and glycine. There is a 50x difference in sensitivity based on subunits and very different timing of sequences.
The part of the receptors inside the cell is the most varied by subunit. These regions determine the signaling into the cell.
Alzheimer’s and Subunits
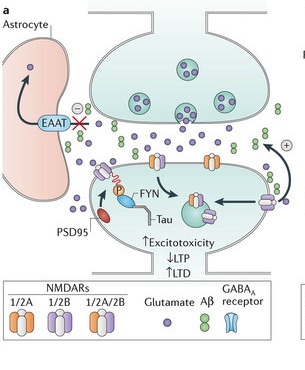 Many subunit effects have been found related to Alzheimer’s.
Many subunit effects have been found related to Alzheimer’s.
- Recently, it was learned that N2B subunits are specifically affected by beta amyloid (one of the potentially destructive agents in Alzheimer’s).
- Also NMDA receptors not at the synapse increase amyloid, which allows N2Bs to impair LTP.
- Tau (another critical factor in Alzheimer’s (which is connected with microtubules) phosphorylates N2B affecting the postsynaptic density increasing excitatory glutamate toxicity.
Another Form of Neuroplasticity by Switching Glutamate NMDA Subunits
There are an increasing number of ways being found that neurons respond to subjective experience. Each type of molecular structure in the brain is responding at once in different ways to the stimulus of subjective mental activity. Each molecular system appears to respond to direction from mind in its own way—somehow, translated into a wide variety of complex chemical languages, such as folding of proteins, alternate RNA splices, changes of electricity, changes of neurotransmitters and receptors and the exchange of different shaped subunits—all occurring in widely connected circuits of neuroplasticity.
 Somehow, each cell knows how to adjust it’s own individual molecular structures as part of a larger circuit which fires in the realm of milliseconds in possibly thousands of different places. Somehow, between the cells in the synapse, many different changes can occur, which are specific to each region.
Somehow, each cell knows how to adjust it’s own individual molecular structures as part of a larger circuit which fires in the realm of milliseconds in possibly thousands of different places. Somehow, between the cells in the synapse, many different changes can occur, which are specific to each region.
We can now add the language of NMDA subunits to the list of other remarkable languages and self-editing processes in the neurons that respond instantaneously to subjective mental experience.