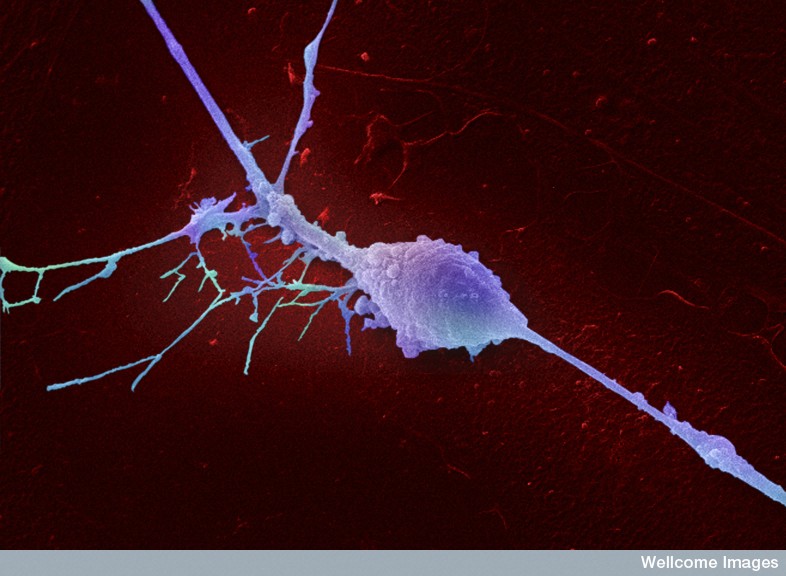
 Some scientists consider scaffolding fibers and tubules in the neuron to be the seat of consciousness. They respond instantly to any mental event with massive movement and construction—building and rebuilding the structures for dendrite spines and axon boutons at synapses in the ever-changing neuron.
Some scientists consider scaffolding fibers and tubules in the neuron to be the seat of consciousness. They respond instantly to any mental event with massive movement and construction—building and rebuilding the structures for dendrite spines and axon boutons at synapses in the ever-changing neuron.
Microtubules are the critical highways for materials, mitochondria and vesicles along the vast length of the axon. Many neuro-degenerative diseases can be traced to dysfunction of microtubules. In fact, the cause of Alzheimer’s might be the disintegration of tau molecules that provide strength and stability to the microtubule structures.
 Scientists are amazed that microscopic materials can be transported more than several feet along one neuron that goes from the spinal cord to the foot. This is equivalent scale to a person carrying a package walking along the wall of China.
Scientists are amazed that microscopic materials can be transported more than several feet along one neuron that goes from the spinal cord to the foot. This is equivalent scale to a person carrying a package walking along the wall of China.
Now, research is showing many different elaborate motors and hundreds of adaptors and factors utilized in this transport. In fact, the extremely complex mechanisms are different for each type of cargo—messenger RNA, small molecules, vesicles filled with neurotrophins, mitochondria, ribosomes, and huge organelles like lysosomes and phagosomes. Each cargo is tagged for its destination. Also, each section of the axon—the initial segment and regions far from the cell body—have different types of transport regulation. Research is showing the enormous complexity of transport along the axon.
Three Types of Tubules and Many Motors for Each
 While there are three basic types of scaffolding tubules—actin, intermediate filaments, and the larger microtubules—it is the microtubules that provide transport along the axon. A previous post discussed these three tubules and another discussed the important myosin motors that work with actin and are critical for neuroplasticity.
While there are three basic types of scaffolding tubules—actin, intermediate filaments, and the larger microtubules—it is the microtubules that provide transport along the axon. A previous post discussed these three tubules and another discussed the important myosin motors that work with actin and are critical for neuroplasticity.
There are thousands of different types of lattice scaffolding structures in human cells—each built with these three molecules. Actin builds a membrane’s moving edge for a growing axon or dendrite. Microfilaments are the most flexible that make strong connections by elaborate branching. Microtubules provide stable structure for transportation.
Microtubules Build Structures Using Many Other Molecules
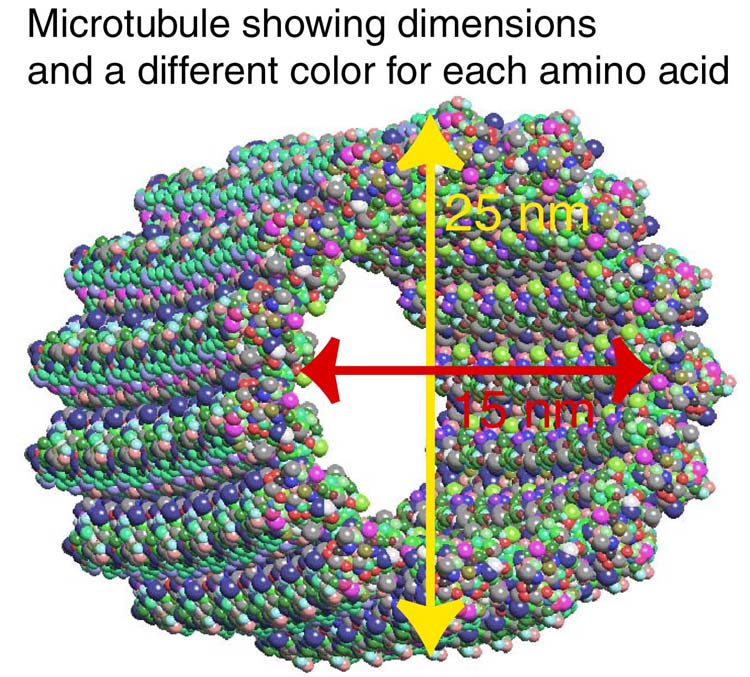 Microtubules are the strongest and most elaborate tubules. They are used for major transportation traffic as well as complicated scaffolding at all levels of neuronal function. They, like the other tubules, are critical for neuroplasticity. Microtubules have hundreds of helper molecules that are needed in different situations.
Microtubules are the strongest and most elaborate tubules. They are used for major transportation traffic as well as complicated scaffolding at all levels of neuronal function. They, like the other tubules, are critical for neuroplasticity. Microtubules have hundreds of helper molecules that are needed in different situations.
The microtubule is built as a spiral cylinder with a positive charge on the growing leading edge and a minus charge on the other. Transport away from the cell body carries lipids, proteins, energy producing mitochondria, vesicles of all types and other materials for the synapse. Transport back to the cell body is critical for mitochondria going back and forth, removal of debris in vesicles and signals related to damage of the distant axon regions. In fact, defects in the ability to transport debris might be the primary cause of Alzheimer’s disorder.
 Microtubules make a reliable shape for a track where motors shuffle or step along carrying many kinds of loads. There are many motors that use these tracks. But, the two major ones in the neuron are the protein motor kinesin, which moves away from the cell center toward the synapse and the protein motor dynein, which moves material toward the cell center.
Microtubules make a reliable shape for a track where motors shuffle or step along carrying many kinds of loads. There are many motors that use these tracks. But, the two major ones in the neuron are the protein motor kinesin, which moves away from the cell center toward the synapse and the protein motor dynein, which moves material toward the cell center.
Rapid growth of axons occurs with many parallel microtubules enlarging at the plus end. The organization of microtubules is much more variable and complex in the smaller, but equally rapidly growing dendrites. For the dendrite, the minus end near the cell body is either connected to the centrosome (an organelle that serves as the main microtubule organizing center) or they are capped for stability.
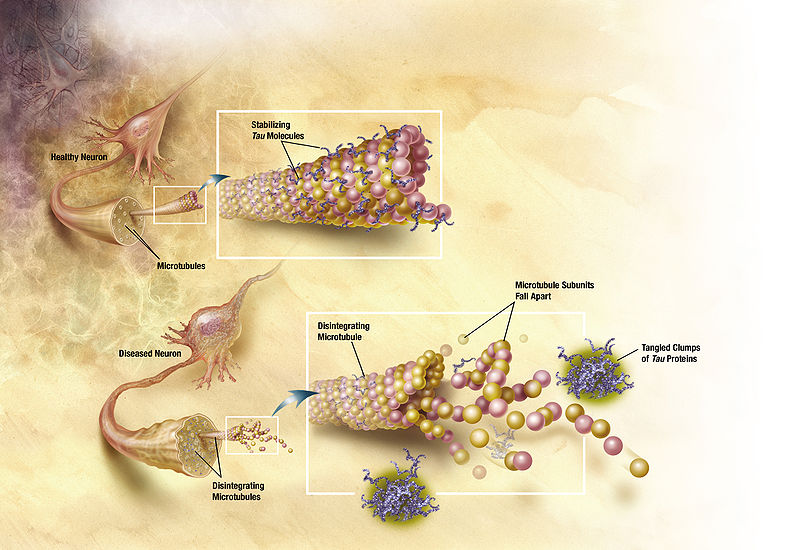 There are a host of associated proteins that maintain stability. A famous one is tau that holds these arrays of microtubules together until they collapse in Alzheimer’s and form neurofibrillary tangles (see post). There are, however, many other proteins associated with microtubules that, when defective, produce disease. These include many different scaffolding proteins, motors and adaptors used to transport different material. These associated proteins can, even, regulate the specific motors on the microtubule.
There are a host of associated proteins that maintain stability. A famous one is tau that holds these arrays of microtubules together until they collapse in Alzheimer’s and form neurofibrillary tangles (see post). There are, however, many other proteins associated with microtubules that, when defective, produce disease. These include many different scaffolding proteins, motors and adaptors used to transport different material. These associated proteins can, even, regulate the specific motors on the microtubule.
There is one group of proteins that interact with the growing positive tip of the microtubule, called +TIPs (plus end interacting proteins). Taking care of the growing tips is a particularly difficult job because tips are unstable and always changing in different ways—growing, rapidly shortening, dramatically breaking and being rescued.
Kinesin and Dynein Motors
Kinesins are the motors that travel from the cell body to the synapse. Dyneins go back to the cell body. They were both discovered thirty years ago and operate with a stepping motion as if they are walking carrying a bundle.
Kinesin Motors
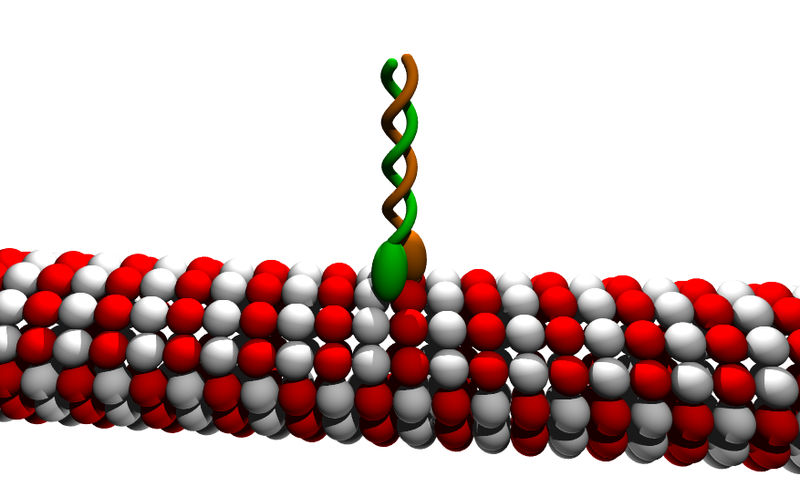
There are 45 genes that regulate and manufacture Kinesin—brain cells utilize 38 of these. The neuron is unique in its complex usage of these motors and their microtubules, so there is a much greater regulation than in other more stable cells. There are many versions of the three basic kinesins—kinesin1, 2 and 3.
Kinesin-1 takes proteins, organelles, RNA and vesicles at a rate of around 1um/second (a micrometer is 0.000001 of a meter). The structure includes two heavy subunits and two light subunits. The light chains are involved in mechanisms for stopping the transport. They move toward the plus growing end of the microtubule. Each step is 8 nm long (a nanometer is 0.000000001 of a meter). This step has a strong attachment and force and can win a match with an opposing motor. But, they still can detach when it is too difficult. New proteins are transported very rapidly. Organelles have been clocked with movement of 400 mm/day (which is 1um/second).
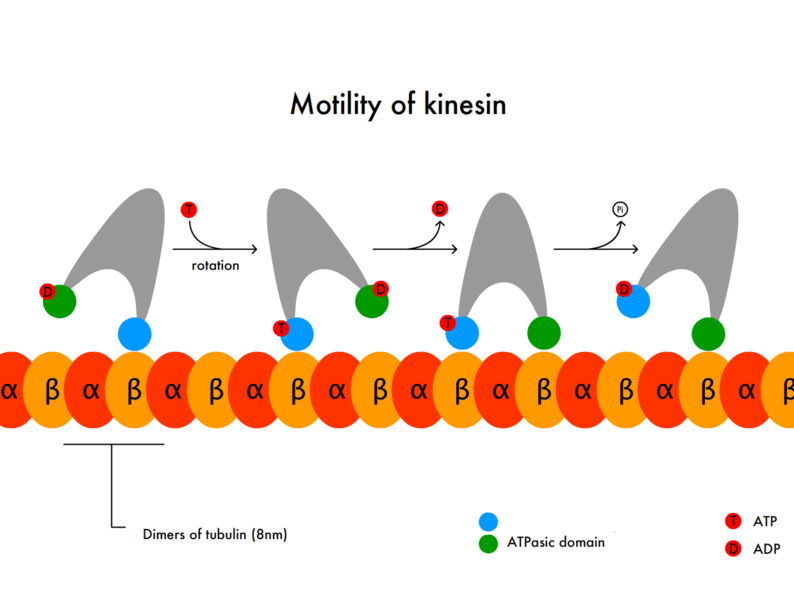 The moving part of Kinesin-2 is either a single molecule or made of multiple subunits. It specializes in carrying material for membranes. The attachment is not as strong as K1 and can detach if presented with a tug or war from an opposing motor.
The moving part of Kinesin-2 is either a single molecule or made of multiple subunits. It specializes in carrying material for membranes. The attachment is not as strong as K1 and can detach if presented with a tug or war from an opposing motor.
Kinesin-3 is altered based on what it is carrying and can be made to travel much faster when carrying critical organelles. K3 carry sacs with neurotransmitters and materials for the synapse. These can operate with increased force because of the importance of their cargo that can overcome opposing transport and obstacles.
Dynein Motors
Unlike kinesin, the active dynein motor subunit is made from one gene. The intermediate and light chains have two genes each. It, also, has heavy and light chains for different structures used in carrying different loads. These subunits can self assemble into different arrangements for different situations.
 Dynein travels very fast, but can, also, take back and side steps, unlike kinesins. It is not as strong as kinesin and loses direct battles with it when traveling in opposite directions along the same microtubule. But, its sideways movement makes it very effective in overcoming obstacles. Dynein can work in teams of motors for larger more complex cargo.
Dynein travels very fast, but can, also, take back and side steps, unlike kinesins. It is not as strong as kinesin and loses direct battles with it when traveling in opposite directions along the same microtubule. But, its sideways movement makes it very effective in overcoming obstacles. Dynein can work in teams of motors for larger more complex cargo.
Dynein, most of the time, needs a factor called dynactin, a very complex molecule made of several proteins that activates dynein for use. It is critical for the function of neurons by binding to the microtubule and the dynein motor. Dynactin has many different binding sites and has a wide range of activity including special functions in the initial axon region and other distant regions.
Motors In Competition – How Are They Regulated
Many cargoes use multiple motors at once, sometimes as many as a dozen. Heavy or large cargoes, such as lysosomes, have many attached motors for transport that work together. These teams of motors can consist of kinesin-1, kinesin-2, and dynein together. Kinesin 1 and 2 work together to transport vesicles with prions inside. Autophagosomes (for cleaning debris) travel a long way on the axon with dynein and kinesin working together.
 How are all these different motors regulated? Is each regulated or is there overall direction; or are the motors competing? One model says kinesin is strongly regulated and less for dynein. However, very new research finds that in many cases, the scaffolding molecules regulate this tug of war. (Another finding that supports consciousness in these molecules).
How are all these different motors regulated? Is each regulated or is there overall direction; or are the motors competing? One model says kinesin is strongly regulated and less for dynein. However, very new research finds that in many cases, the scaffolding molecules regulate this tug of war. (Another finding that supports consciousness in these molecules).
Each type of cargo, motor and adaptor seems to be regulated differently. There is reason to think that for lysosomes there might be a tug of war between kinesin and dynein. For autophagosomes the dynein is dynamically lowered by the microtubule so that kinesin can push it forward.
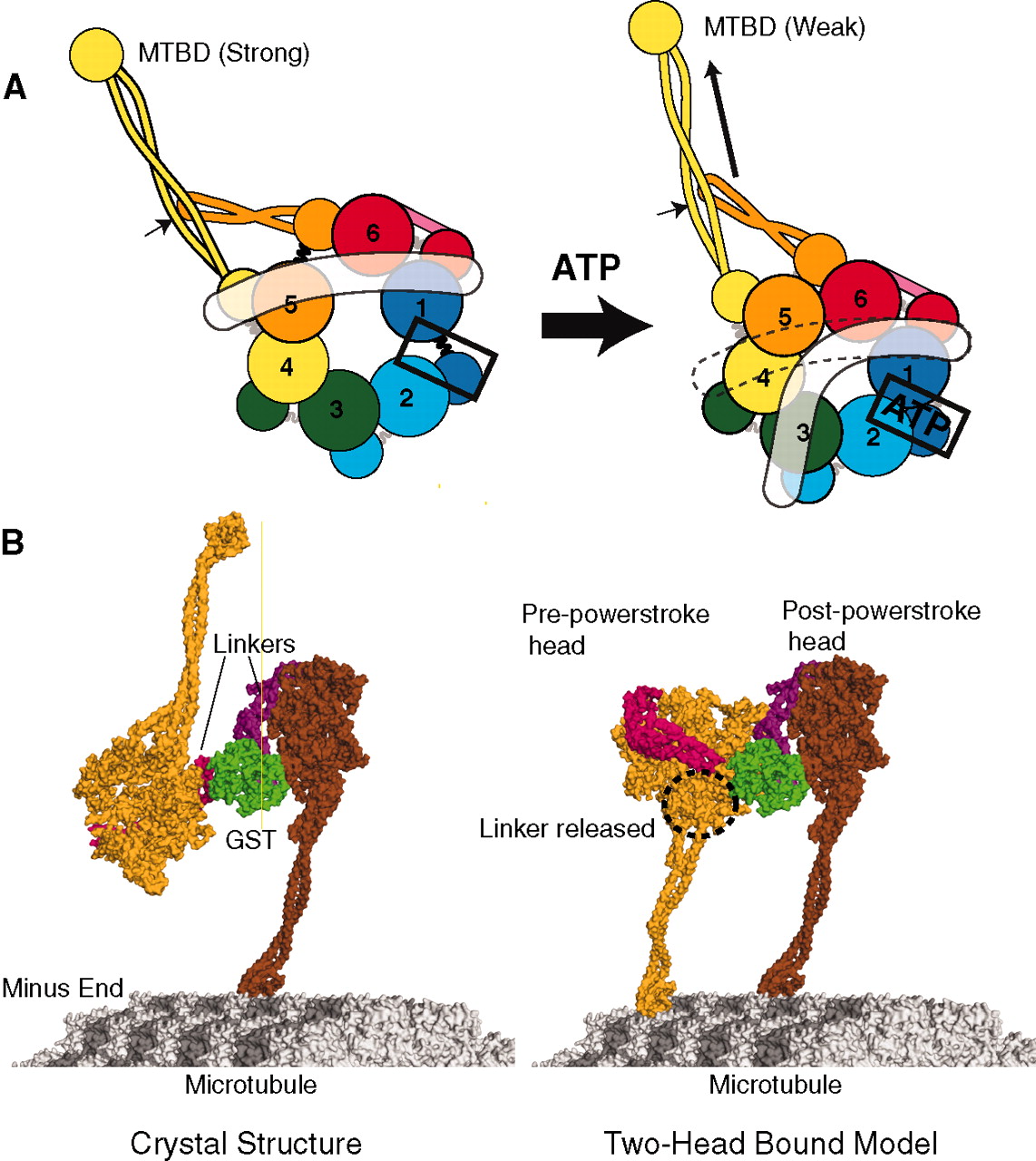 Kinesein-1 has been shown, at times, to regulate itself and change its speed and force. The inhibition occurs with factors that bind to kinesin’s tail where it connects with the motor. Scaffolding proteins alter this inhibition, which can cause movement in either direction.
Kinesein-1 has been shown, at times, to regulate itself and change its speed and force. The inhibition occurs with factors that bind to kinesin’s tail where it connects with the motor. Scaffolding proteins alter this inhibition, which can cause movement in either direction.
There are, in fact, many factors that alter movement in different ways with different cargoes. Dynein regulation can be caused by attachment and detachment of ATP energy molecules, which alters its force. This causes the motor to attach more strongly to the microtubule with less movement. Another factor, the protein huntingtin, regulates vesicles with BDNF and autophagosomes, and is relevant to Huntington’s disease. There are many enzymes involved in this process.
Transport Regulation Mechanisms Can Be General or Specific
Each large organelle takes part in the regulation of its own movement and they use quite different mechanisms. For some organelles, specific motors stay with an organelle even if the motor is not being used at that time. Specific groups of opposing motors move very large organelles. These mechanisms are very complex and include kinase enzymes, multiple molecular cascades, attaching phosphorus energy particles, and scaffolding protein activity.
Fast Forward Transport for Neurotrophins
Neuropeptides, neurotransmitters, and neurotrophins are critical to neuron function and are carried in special vesicles called dense core or granular vesicles by kinesin-3. The mechanism for granular vesicles changes the structure of the kinesin-3 subunits as well as the specific adaptors. These particular vesicles cannot be returned and new ones must be sent all the way from the cell body to the synapses. Each is tagged for delivery to specific dendrites and axon tips with different material. There are specific regulators of the movement as it approaches the target. The dynein is disabled at the site to avoid return of useless sacs. The sacs are recycled locally.
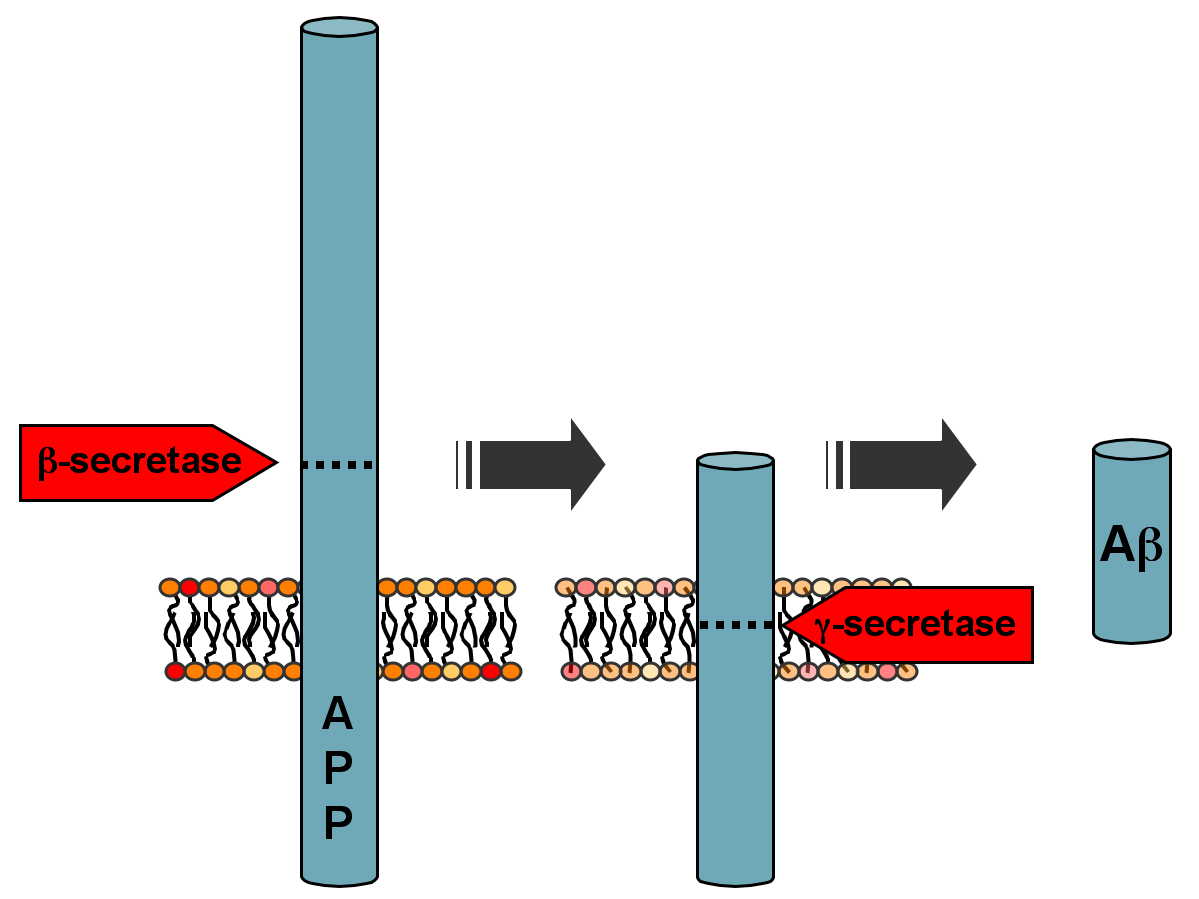 Protein that is the precursor of amyloid, APP (amyloid precursor protein), is carried in a vesicle at very rapid rates mostly away from the cell body. The mechanism includes a particular molecule that is phosphorylated to provide the switch determining direction.
Protein that is the precursor of amyloid, APP (amyloid precursor protein), is carried in a vesicle at very rapid rates mostly away from the cell body. The mechanism includes a particular molecule that is phosphorylated to provide the switch determining direction.
BDNF (brain derived neurotropic factor) is a critical molecule for neuronal function and is transported from the cell body to the synapse. The vesicle transport, uniquely, uses huntingtin and makes a scaffolding structure with kinesin-1 and dynein motors. Huntingtin phosphorylation is the switch to go forward and the opposite dephorphorylation goes backward. (A defective huntingtin causes Hungtington’s disease)
Neurotrophins are factors that are secreted locally near the synapse and maintain the health and very life of the neuron. Without them the neuron will die. These critical molecules bind at the presynaptic terminal–the end of the axon. From there, they are transported backwards to the cell body where they stimulate networks of genes to maintain the health of the neuron. It is a signal from the tissue where the axon has landed to help keep the neuron alive.
 Neurotrophins are taken into the cell in vesicles (signaling endosomes) and taken along the microtubules to the cell body. This transport is very active, fueled by ATP production along the axon from mitochondria. Mitochondria are moved (see post on microtubules alive in the cell) along the microtubules to provide this needed energy (30% of mitochondria are moving at any one time).
Neurotrophins are taken into the cell in vesicles (signaling endosomes) and taken along the microtubules to the cell body. This transport is very active, fueled by ATP production along the axon from mitochondria. Mitochondria are moved (see post on microtubules alive in the cell) along the microtubules to provide this needed energy (30% of mitochondria are moving at any one time).
Transporting Mitochondria for Energy
 The movement of mitochondria has unique regulation. Mitochondria are slowed near the synapse so they will stay there and provide energy. The signal to slow is the increased levels of calcium from the neuron’s action potential at very particular places near the synapse.
The movement of mitochondria has unique regulation. Mitochondria are slowed near the synapse so they will stay there and provide energy. The signal to slow is the increased levels of calcium from the neuron’s action potential at very particular places near the synapse.
Those mitochondria that are nearby–15nm away–are not affected. But, if they travel through the area of increased calcium they stop. When specific dendrites are very active, the mitochondria maintain a higher level of energy production. There are many complex mechanisms of the transport motors and adaptors to provide this calcium effect.
 In previous posts, microbes’ decision-making ability, communication, and group behavior were discussed. It described the process of mitochondrial fission and fusion that is highly regulated through the endoplasmic reticulum of the neuron. Mitochondria within the neuron function as a microbe community working together for the goal of providing energy throughout the very complex neuron. They communicate with each with various signals and to the cell through intermittent contact with the endoplasmic reticulum.
In previous posts, microbes’ decision-making ability, communication, and group behavior were discussed. It described the process of mitochondrial fission and fusion that is highly regulated through the endoplasmic reticulum of the neuron. Mitochondria within the neuron function as a microbe community working together for the goal of providing energy throughout the very complex neuron. They communicate with each with various signals and to the cell through intermittent contact with the endoplasmic reticulum.
Transporting Endosomes and Lysosomes
Critical vesicles, such as endosomes, phagosomes and lysosomes, are carried on the transport systems, many going back and forth rapidly. Like mitochondria these are essential organelles that are needed to clean up debris or bring information to different regions. There are unique mechanisms for the transport away from the cell body using adaptors that link to the subunits of kinesin-1 and 2, and others to dynein.
Transport of Structural Proteins for Scaffolding
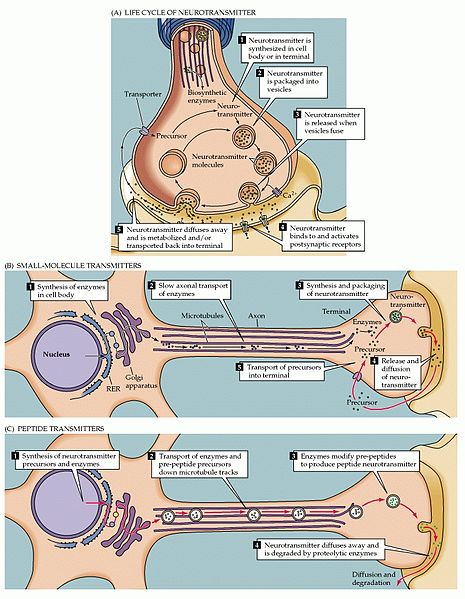 The transport of hundreds of different structural materials to constantly build and rebuild the structure of the axon is slower than that of the organelles. There are two different slow speeds for these materials—1 mm per day and 10 mm per day. The slower speed is for intermediate filaments and tubulin (to make microtubules). The faster rate is for a large number of smaller molecules. It is much harder for scientists to study the slow transport because of the need for very long observation of living microscopic material.
The transport of hundreds of different structural materials to constantly build and rebuild the structure of the axon is slower than that of the organelles. There are two different slow speeds for these materials—1 mm per day and 10 mm per day. The slower speed is for intermediate filaments and tubulin (to make microtubules). The faster rate is for a large number of smaller molecules. It is much harder for scientists to study the slow transport because of the need for very long observation of living microscopic material.
Different intermediate filament assembled units are transported by kineisin-1 and dynein. One of the subunits binds directly to the motors. The transport seems to be in small movements with rests in between. The transport of actin and microtubule structures is not as well known. It is especially difficult to study microtubules, because they are built up and broken down so rapidly. Tubulin for microtubules is transported either as two molecules or as small microtubule assemblies.
More than two hundred other molecules are transported to be used in construction. Synapsin is one critical protein that combines into complexes, which latch onto passing motors. The slow rate of transport is related to the complex attaching and reattaching with periods of travel and waiting. Despite this type of stop and go system, the slow transport provides three times the material as the fast transit. The proteins that travel from the spinal cord to the foot can take from four months to a year for transport.
Transport Control in Different Axon Regions
 Each region of the axon has very different scaffolding structures and different mechanisms.
Each region of the axon has very different scaffolding structures and different mechanisms.
In the initial segment there is a very specific structure with unique stabilizing molecules. This region makes sure that cargoes headed for the dendrites and the cell body don’t start travelling down the long axon. Some cargoes are targeted for the dendrite, but start down the axon and are stopped and re routed. Unique adaptors and modifiers for microtubules in this region help this process.
The distant regions of the axons direct the return of cargoes all along the axon to the cell body by unique mechanisms. The growth of microtubules in these distant regions is different. It is much more active and variable with many different plus ends appearing. There are special proteins in this region that interact with the active plus ends. Dynactin, also, interacts with the special proteins at the plus ends. These complex very active mechanisms appear to give direction to transport heading back to the cell body
Since the electrical charge seems to be related to this increased intelligent activity of the microtubules, it is has be questioned whether electrical fields are involved. Please see the post on electrical fields and gradients determining cellular behavior.
Making Proteins All Along the Axon
 While many of the proteins are manufactured in the cell body and transported the long way to the synapse, there is increasing evidence that there is, also, local ribosome activity all along the axon and in the dendrites. Large ribosomes are, first, transported to distant sites. To use the local ribosomes, messenger RNA is transported to these regions.
While many of the proteins are manufactured in the cell body and transported the long way to the synapse, there is increasing evidence that there is, also, local ribosome activity all along the axon and in the dendrites. Large ribosomes are, first, transported to distant sites. To use the local ribosomes, messenger RNA is transported to these regions.
Research has uncovered more about the production of local proteins in the dendrite. In order to transport messenger RNA they need to be altered. The usual mechanisms to find a ribosome are held in check until they travel to the distant ribosome. Specially altered messenger RNAs are put in a granule vesicle with the necessary proteins and even some large ribosomes. These specially packed vesicles go back and forth in both directions—wherever needed—and seem at times to oscillate.
The movement of messenger RNA vesicles is greatly increased when there is nerve injury, as well as by some chemical signals. When there is injury, special proteins are manufactured that bind to the motors stimulating more rapid transport of supplies. There are, also, genetic activating factors that stimulate more protein manufacturing. With nerve injury, complex multi protein structures produce transport back to the cell. They signal the activation of genetic machinery to rev up manufacturing for repair. This process involves many steps from a variety of enzymes triggered by energy rich phosphorus.
 A faster signal of the injury is increased calcium at the injury site that radiates back to the cell body. This process affects the histone regulating enzymes in the nucleus causing more genetic activity to make needed materials for transport. Chemical signals can, also, increase transport of more messenger RNA vesicles. Special factors, like NGF, can signal production of proteins from messenger RNAs in distant sites.
A faster signal of the injury is increased calcium at the injury site that radiates back to the cell body. This process affects the histone regulating enzymes in the nucleus causing more genetic activity to make needed materials for transport. Chemical signals can, also, increase transport of more messenger RNA vesicles. Special factors, like NGF, can signal production of proteins from messenger RNAs in distant sites.
Surprisingly, even before injury, vesicles filled with messenger RNA are placed along the axon, ready to be used when needed.
Transport Needs Energy
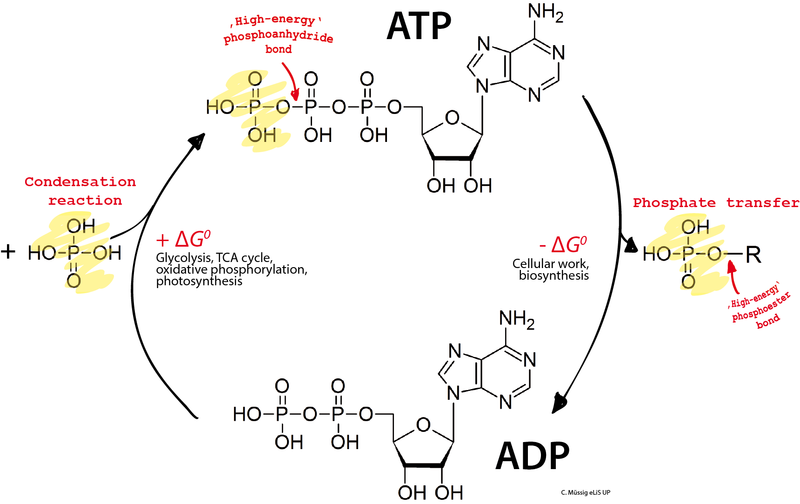
The transport motors stepping along the axon use ATP as their energy source. Kinesin-1 uses one ATP for every step of 8 nm. Each large vesicle or organelle has multiple motors walking. If there are conflicts and tug of wars, the amount of energy is increased. A typical transit can use millions of energy molecules. Dynein’s steps are much larger, up to 32nm, but it can go backwards and sideways as well as forward. Dynein uses many motors per vesicle—up to a dozen. This process, also uses a million energy molecules.
Given the large amount of energy required for axon transport, it is not surprising that a second mechanism has been discovered unique to transport. While mitochondria probably supply most of the energy, it appears that vesicles have glycolytic enzymes that produce energy particles linked to the vesicles as they travel on the microtubules.
 In fact, the energy needs of neuronal baseline activity are very high and transport by motors is not a great percentage of the total. The brain, weighing three pounds, uses at least 20% of all the energy of the body. Much of the usage is the constant electrical activity along the axons, which requires maintenance of a large number of membrane channels. The vesicle energy backup system could work in the regions between mitochondria. This type of energy can only be used for the larger cargoes with vesicles.
In fact, the energy needs of neuronal baseline activity are very high and transport by motors is not a great percentage of the total. The brain, weighing three pounds, uses at least 20% of all the energy of the body. Much of the usage is the constant electrical activity along the axons, which requires maintenance of a large number of membrane channels. The vesicle energy backup system could work in the regions between mitochondria. This type of energy can only be used for the larger cargoes with vesicles.
Another important factor is the energy support from glia cells. Glia provide materials for glycolysis as well.
The Enormous Complexity of Transport Along the Axon
A mental event triggers instantaneous building and tearing down of scaffolding molecules. The scaffolding molecules are critical for every function of the neuron, including rapid changes of neuroplasticity. Somehow, they are the circuit boards and Lego blocks of information flow in the neuron. It is hard to imagine how this direction can occur or where it is coming from.
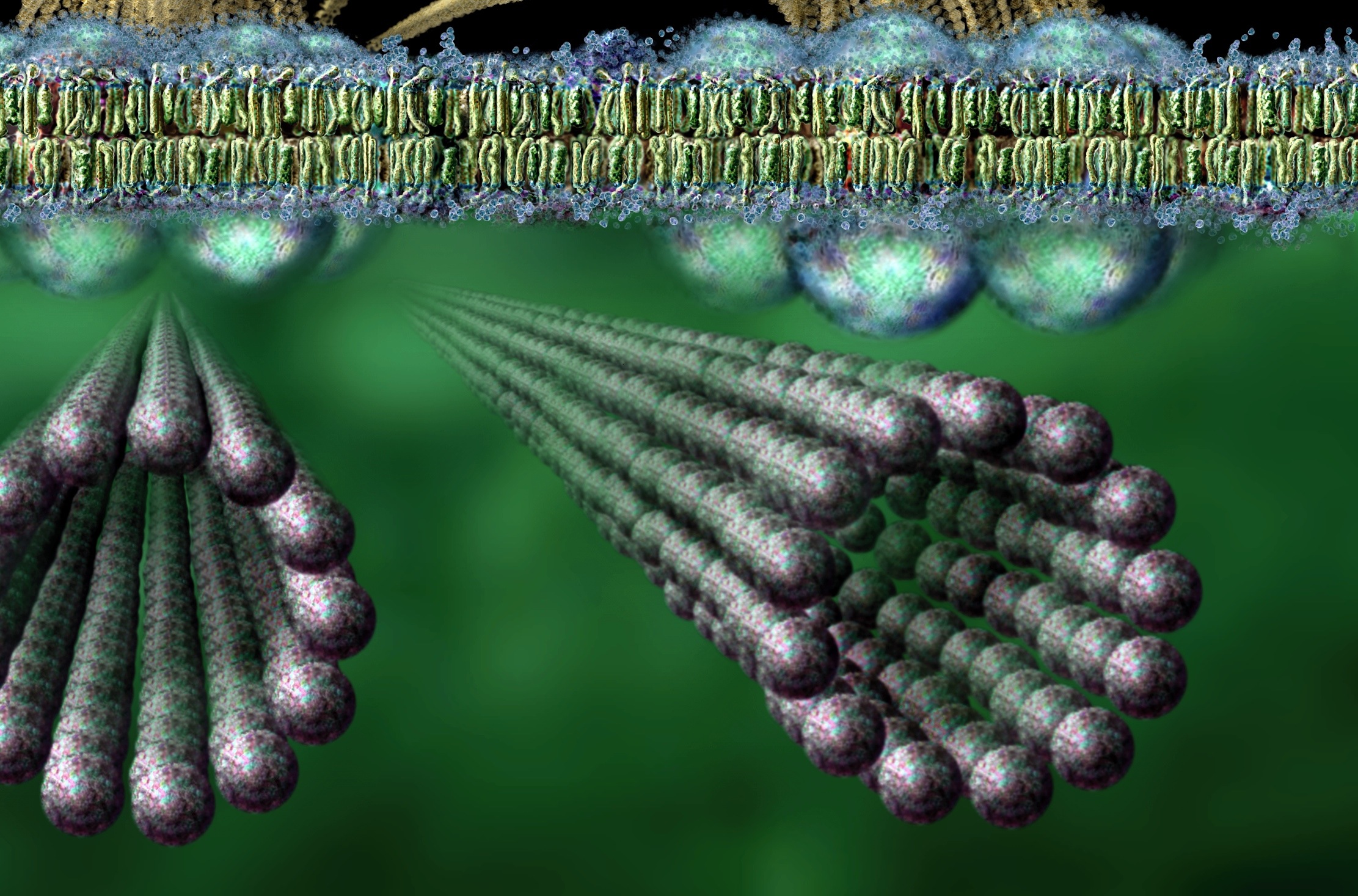
Because of this, some scientists suspect there might be quantum computer activity in the middle of the microtubule. This information at the quantum scale would communicate with other tubules and direct the microtubules and other molecules. Such a scenario, also, fits a theory of mind interacting with quantum states.
In any case, something must direct these massively complex, shifting structures. Does a neuron think with its microtubules? How does this process connect with human thinking?
The transport microtubule system is extremely complicated, where very specific multiple motors, factors and adaptors operate together to transport many different kinds of cargoes. The complexity of the transport system includes tagging and choosing each molecule for specific cargos and specific destinations, while choosing multiple unique co factors and adaptors. Totally different assemblies are chosen for building materials, neurotrophins, mitochondria, and large vesicles like lysosomes. Each uses multiple motors at a time. It is difficult to imagine the direction for all these processes.
With all of this complex activity and regulation responding instantly to thought and the alterations of neuroplasticity, how can anyone think that this process is in any way random?