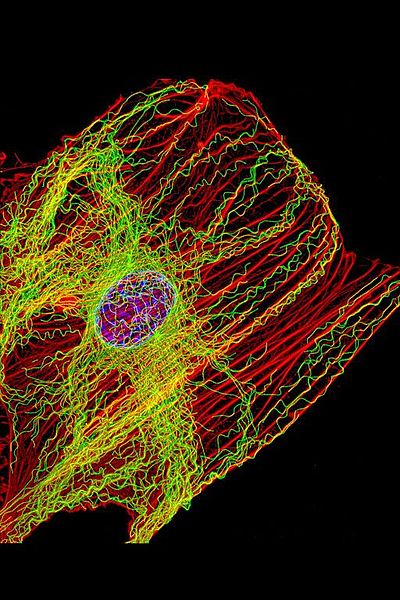
 Microtubules may be the brains of the cell, particularly neurons—operating like a computerized Lego set. They are large complex scaffolding molecules that work closely with the two other rapidly changing structural molecules, actin and intermediate filaments, to provide structure for the entire cell including the spatial placement of organelles. In neurons, microtubules respond instantly to mental events and constantly build and take down elaborate structures for the rapidly changing axons and dendrites of the synapses. Some think that microtubules are quantum computers and the seat of consciousness. Their lifestyle is quite remarkable. A previous post described elaborate functions along the neuron’s axon including special tagging of cargoes that are transported by distinct motors with complex ancillary molecules for each type of transport.
Microtubules may be the brains of the cell, particularly neurons—operating like a computerized Lego set. They are large complex scaffolding molecules that work closely with the two other rapidly changing structural molecules, actin and intermediate filaments, to provide structure for the entire cell including the spatial placement of organelles. In neurons, microtubules respond instantly to mental events and constantly build and take down elaborate structures for the rapidly changing axons and dendrites of the synapses. Some think that microtubules are quantum computers and the seat of consciousness. Their lifestyle is quite remarkable. A previous post described elaborate functions along the neuron’s axon including special tagging of cargoes that are transported by distinct motors with complex ancillary molecules for each type of transport.
 Microtubules are the basic structural elements for cell division. The centromere is a key structure holding chromosomes together. It connects with the kinetochore where microtubule based spindle fibers attach to the chromosomes. Centrioles produce microtubules that orchestrate the rearrangement and sorting of the DNA during the extremely elaborate process of cell division. Complex arrangements of microtubules direct and pull all the elements of the division process through multiple phases. The structure for this process is considered the most complex machine ever discovered in nature and is based on microtubule actions.
Microtubules are the basic structural elements for cell division. The centromere is a key structure holding chromosomes together. It connects with the kinetochore where microtubule based spindle fibers attach to the chromosomes. Centrioles produce microtubules that orchestrate the rearrangement and sorting of the DNA during the extremely elaborate process of cell division. Complex arrangements of microtubules direct and pull all the elements of the division process through multiple phases. The structure for this process is considered the most complex machine ever discovered in nature and is based on microtubule actions.
Microtubules are critical for the neuron’s migration; for the polar structure of the dendrite, soma and axon; and for stem cell’s determination of the type of cell a neuron will become. They are highways for long distance transport of materials and organelles. They are dynamic and changeable with elaborate mechanical functions. They control the signaling at local regions of the extremely long axon.
The control of the microtubule function is extremely complex, establishing and maintaining the architecture of the neuron. The control is distributed throughout the cell, which brings up the question of where the central control is to respond to mental events in such detailed and complex ways. They are highly regulated in terms of the number, length, allocation, exact positioning and placement. Abnormalities in any of these functions leads to brain disease. Mutations in the proteins that hold the microtubules in place, such as tau are critical for the development of major neurodegenerative diseases (see post). The motors that move material along the microtubule highways—dyneins and kinesins—and regulators of these motors—dynactins—can produce other brain diseases. Defective transport in the axon is associated with ALS, Alzheimer’s, and multiple sclerosis.
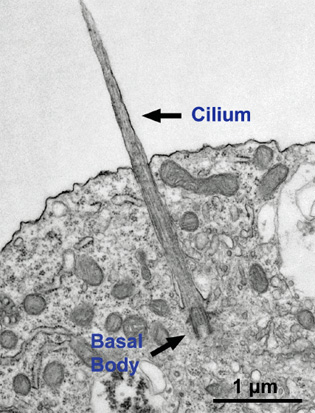 Neurons experience major shape changes as they move into place and when they build axons, axon-boutons (endings), dendrites with spines, and elaborate synapses. All of these are dependent on microtubules working with the constantly changing actin cytoskeleton. They set up signals for the placement of compartments and organelles in the cell. They are involved in signaling and the critical elements of the primary cilium, noted in a previous post to be the antennae and center of all of the cell’s activity.
Neurons experience major shape changes as they move into place and when they build axons, axon-boutons (endings), dendrites with spines, and elaborate synapses. All of these are dependent on microtubules working with the constantly changing actin cytoskeleton. They set up signals for the placement of compartments and organelles in the cell. They are involved in signaling and the critical elements of the primary cilium, noted in a previous post to be the antennae and center of all of the cell’s activity.
The microtubule structures are so complex that many of the mechanisms are not yet known. Recent research tools with extreme resolution have shown that these mechanisms are much more complex than previously thought. Signaling pathways have been found that regulate the building, maintaining and rebuilding of microtubule structures.
Microtubule Basics

There are many versions of 7 kinds of tubulin molecules named α, β, γ, δ, ε and ζ. The basic linear proto filament is a structure built from two kinds of the protein molecule tubulin—α-tubulin and β-tubulin—put together head to tail and then organized into a hollow tube approximately 25 nm in size. To start structures, a third type of γ-tubulin is necessary. γ-tubulin combines with other large proteins to form a ring complex that is the starting place for the structures. This origination is called nucleation.
The structures are built and then taken apart constantly, while the microtubule grows into new regions and then pulls back when the situation changes or the environment is not correct for the structure that is being built. The building and un building is based on the energy source of GTP which binds with free tubulin. One view is that GTP forms a cap at the growing end to hold the structure together. If the GTP breaks apart the structure breaks apart.
The two different ends of the growing hollow tube are different. The plus end grows rapidly and just as rapidly can break apart. There are special proteins that grow with the tube and regulate some of the process—tracking proteins or +TIPS. Other proteins are called end-binding proteins EB that connect with many other structures such as motors and actin structures.
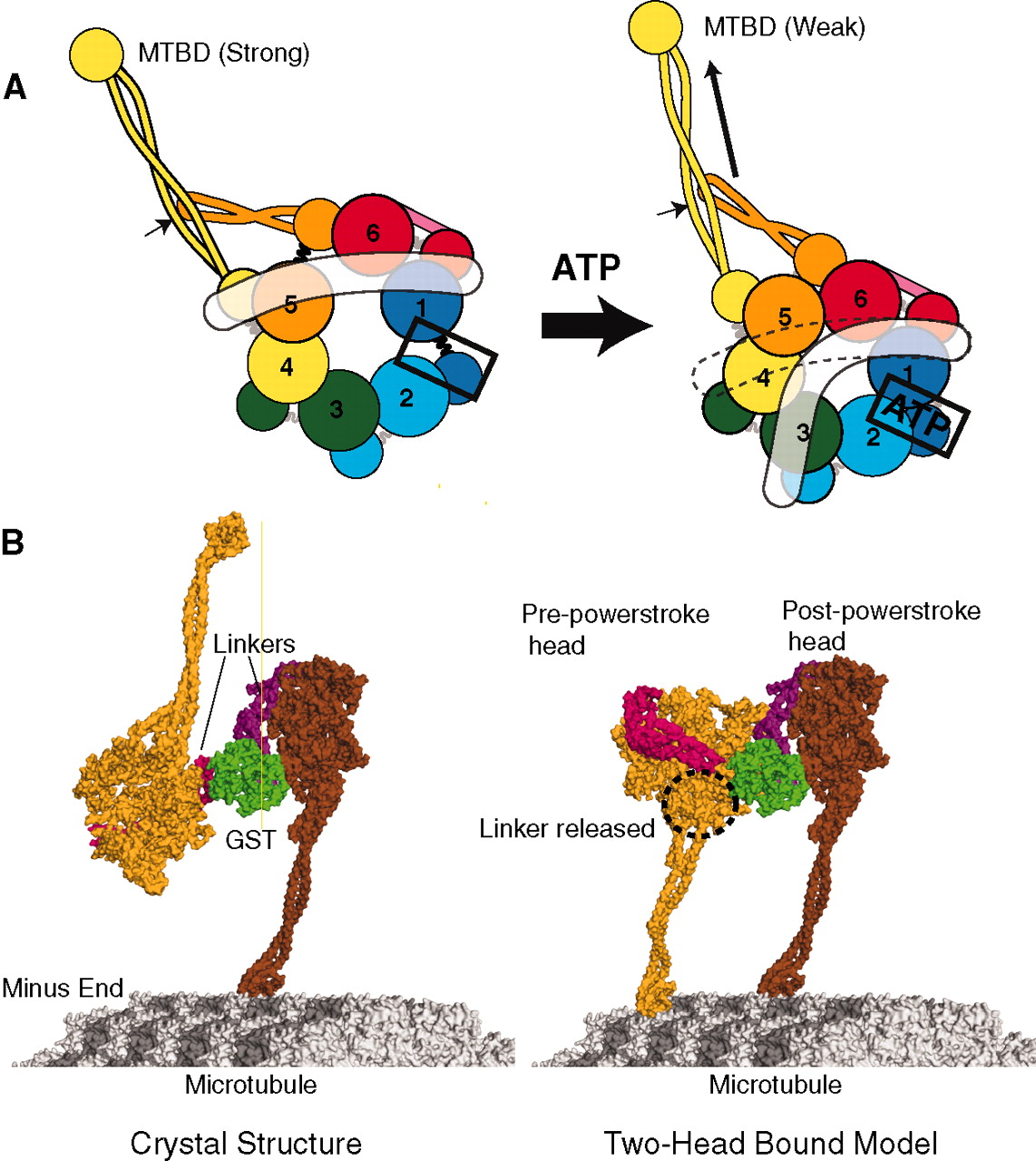 In fact, it is much more complex. The assembly has many other necessary complex factors including microtubule associated proteins MAPs, other tubulin isotypes, motor proteins and motor attachment proteins, and PTMs or post translational modifications.
In fact, it is much more complex. The assembly has many other necessary complex factors including microtubule associated proteins MAPs, other tubulin isotypes, motor proteins and motor attachment proteins, and PTMs or post translational modifications.
There appear to be at least five different types of associated proteins. One binds and regulates the dynamics such as + and – TIPS. A second type creates cross-links and stabilization of structures. Another type regulates the sites of nucleation and where the structures are started. This group also breaks apart structures. Another group are motors, such as kinesin and dynein that create movement and mechanical forces related to building structures. The fifth are special proteins that influence the folding of the tubulin molecules and modify the structures. This last group creates many different types of unique structures.
Materials are Tagged for Transport on Microtubule
 One of the major functions of microtubules is to regulate all transport along the very long axon as well as the cell body and dendrites with unique spines. Specific material has to be sent to each area. Cells are very small compared to humans—the size of a person compared with Mount Everest. However, when looking at the scale of neurons, they can have axons that are multiple feet long (from spinal cord to hands or legs). Transport at this scale is that of a human walking along the wall of China.
One of the major functions of microtubules is to regulate all transport along the very long axon as well as the cell body and dendrites with unique spines. Specific material has to be sent to each area. Cells are very small compared to humans—the size of a person compared with Mount Everest. However, when looking at the scale of neurons, they can have axons that are multiple feet long (from spinal cord to hands or legs). Transport at this scale is that of a human walking along the wall of China.
The neuron must send a wide variety of specific tagged material to particular places in the cell and along the axon. There is a great amount of signaling regulating this transport. The motors carry organelles, molecules to make vesicles, protein receptors, molecules that are involved in migration of the axon and microRNAs. See the post on Complexity in Transport Along the Axon, for detailed description of how material is tagged, loaded, anchored and delivered. Different kinds of tubules exist for axons and dendrites (+ to the axon and – to the dendrites.) Special motors exist for each.
Microtubule Structures Shape the Entire Cell
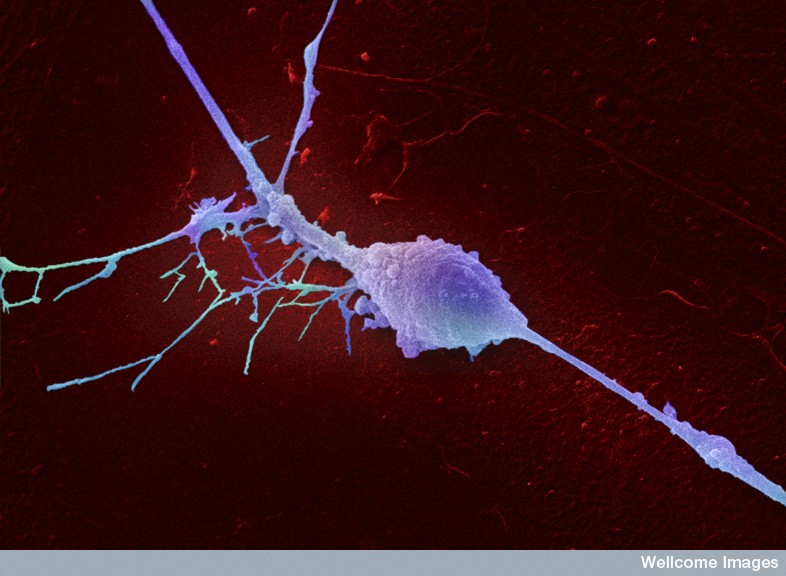
When a neuron is being produced and while it migrates, it changes shape. Other shape changes occur frequently with new synapses both axons and dendrite spines (see post on Dendrite Spine Complexity). When a neuron migrates it produces a process in front, moves the nucleus to the front and then disassembles the process left in back. Microtubules and actin scaffolding direct all of this.
The anchor in this process is the centrosome made of centrioles that are made of specific microtubules structures. It produces microtubule connections in the processes moving forward. The centrosome is the organizing center of the actions of microtubules. It is an organelle near the nucleus. Two centrioles at right angles are surrounded by a large protein mass. This very complex machine and directs cell division by pulling all elements of the division through many phases.
The centriole is a set of microtubules often arranged as a tubule with nine microtubules defining the circle. In some there are other microtubules in the center. (See post on the Primary Cilium for a discussion its centrioles). When centrioles connect, they do so at right angles and these pairs travel to opposite ends of the nucleus in the cell division process. Centrioles are involved in the organization of the mitotic spindle and in the completion of cytokinesis, or movement of the DNA in cell division.

But, centrosomes, made of centrioles, are, also, the critical way the neuron organizes the spreading and constantly changing microtubule structure. In fact, the centriole determines where the nucleus sits in the cell as well as organizing spatial structure of organelles in the cell (golgi, endoplasmic reticulum, etc). In cells with cilia and flagella, the central centriole determines where it will be. This mother centriole is, also, called the basal body as the starting point of the cell’s entire microtubule process.
Microtubules make a large structure that surrounds the entire nucleus in a cage. This cage extends from the centrosome around the nucleus and into the leading process. These microtubules orchestrate the neuron’s ability to migrate. The tubule structure then pulls the centrosome with the nucleus into the leading edge.
 When the axon starts and grows the cell’s shape becomes polar and asymmetric. The neurite (start of the axon) grows with bundles of microtubules and a very active actin growth cone. This complex process involves mechanical actions of both.
When the axon starts and grows the cell’s shape becomes polar and asymmetric. The neurite (start of the axon) grows with bundles of microtubules and a very active actin growth cone. This complex process involves mechanical actions of both.
When the neuron becomes a specific type, microtubules take on very specific shapes and have to maintain these with unique stabilizing molecules. This involves very active transport of these stabilizing molecules by kinesin motors. How this is directed is not clear. It is possible that the centrosome and Golgi are involved. As the axon grows longer, many different stabilizing proteins are involved, such as +TIPs, interacting with actin, adhesion molecules, and the microtubules. At times an entire bundle of many microtubules are moved by mechanical forces from motors to allow for changes in shape. When injury to the axon occurs, microtubules are again critically involved in the rebuilding.
The microtubules have many different roles in forming and stabilizing synapses. A previous post showed the dynamic changes of dendrite spines and the different shapes. This occurs through microtubule actions. The growth into the spines is regulated by factors like BDNF. These microtubules bring material for the changing spine shapes with special motors.
Organization and Structure of Cytoskeleton
 Axons can have as many as 100 bundles of microtubules in one axon cross section. There are many variations in these lattices with different types of stabilizing molecules, different orientations, and many different associated molecules and co factors. It is so complex that much of the structure is not understood despite extensive research with electron microscopes and thin sections.
Axons can have as many as 100 bundles of microtubules in one axon cross section. There are many variations in these lattices with different types of stabilizing molecules, different orientations, and many different associated molecules and co factors. It is so complex that much of the structure is not understood despite extensive research with electron microscopes and thin sections.
The initiation and organization of the microtubule structure is often directed by the large MTOC (microtubule organizing center) with the centrosome as one example. They can generate from the Golgi and even in distant regions along other existing tubule structures. The minus ends are therefore not always at the centrosome. The very first structures start in the centrosome but then as it gets more complex and larger throughout the axon this direction appears to fade and others take it up. A previous post described the critical cellular cilium with their many functions in signaling and movement. These cilia are highly organized by microtubules of particular shape (9 plus 2) and they originate from the centrosome.
But most of the microtubules are not anchored at all at either end. Orientations are different, also, when starting from different sources. In dendrites the orientation of plus and minus is half and half, whereas in the axon it is mostly + as the lead. Motors such as dynein can use this by going retrograde.
Microtubules are constantly enlarging and shrinking in both axons and dendrites even in a mature axon synapse. It appears that some are stable in these mature situations and others are not. Areas that are more stable have many more associated proteins and linkages.
Types of Microtubule Structures
 There are a variety of different tubulin molecules that build the microtubule and the basic structural α-tubulin and β-tubulin have variants making it more complex. These variants are called isoforms and are produced by different genes, different alterations that occur in the protein once it is manufactured and different filament structure. One difference is the sequence of amino acids in a section of the molecule that sticks out from the structure as a tail in different shapes that form a pattern and a code. These differences in sequence appear to have functions in different types of cells. Mutations in these tails are related to brain diseases. One type has been associated with ALS.
There are a variety of different tubulin molecules that build the microtubule and the basic structural α-tubulin and β-tubulin have variants making it more complex. These variants are called isoforms and are produced by different genes, different alterations that occur in the protein once it is manufactured and different filament structure. One difference is the sequence of amino acids in a section of the molecule that sticks out from the structure as a tail in different shapes that form a pattern and a code. These differences in sequence appear to have functions in different types of cells. Mutations in these tails are related to brain diseases. One type has been associated with ALS.
There are, also, special chaperone molecules that help the tubulin protein molecule in folding. One particular mutation in the chaperone causes a devastating human disease with severe developmental symptoms.
The modifications of the tubulin tails can occur after it is part of the lattice. Some of these modifications can help with the stability of the structure over time. They can attract special molecules that stabilize the structure and stop decomposing of the tubule. There are many modifications of these tails including removing an amino acid, cutting of a section and tagging with acetyl, phosphorylation, glycylation and polyglutamines. There are specific enzymes that work with these tags for particular purposes. Modifications are noted in certain sections of the neurons that obviously have a particular function. This appears to be another elaborate code that is not yet understood. Enzymes with other functions appear to operate on microtubule tails. For example the enzymes that taking off acetyl marks on histones—histone deacetylase or HDAC—also operate on these tails.
The initial segment of the neuron organizes the flow of material into the axon, not allowing diffusion of many proteins that remain in the cell body. It allows certain types of transport into the axon and not others. An unusual bundle of several microtubules are found in this region, which may be related to initiating the action potential. These have many cross-links into a structure called a fascicle. These also appear to be related to regulating the flow of the tau molecule between the axon and the cell body.
Shaping the Microtubule Structure
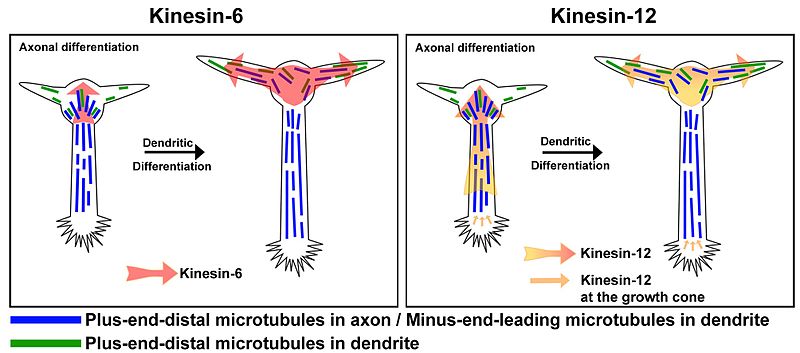
Many different factors, motors, and protein complexes regulate the complex 3D dynamic lattice of microtubules. γ-tubulin forms an elaborate complex to start the process that becomes a template to build the structure at the beginning. It can start in the centrosome or not. It was thought that these non-centrosome structures were cut from the original complex, but there is no real evidence for this. Some organisms have active lattices without any centrosome at all. The original centrosome is taken apart after differentiation of the neuron (choosing a complex shape). Recently some γ-tubulin has been found in axons and dendrites. Origination sites have potentially been found at the Golgi, at the plasma membrane and other places.
Golgi establishes its own elaborate microtubule lattice sending material towards the front of the moving neuron. Golgi appears to have the mechanism to start structures related to other purposes. Golgi has its main operations in the body of the cell but has other outposts in some dendrites that help create the dendrite shapes. But, there appears to be other γ-tubulin present and other sources to start the scaffolding. New lattices can branch off of existing ones as well. Special proteins have been found that binds to the microtubules and then attracts γ-tubulin to start another scaffold.
Special enzymes cut part of a microtubule lattice and use it to create a new lattice. There are three families of enzymes that provide this service—katanin, spastin, and fidgetin—that are part of a large group of enzymes that take apart protein structures. These enzymes appear to be particularly important in making branches in the axon to form multiple boutons and in dendrites making multiple spines. The cuts are not random and are regulated by the special proteins associated with the microtubule structure. They are, also, active during synapse pruning.
One hypothesis is that these cutting enzymes are connected with the defects in the stabilizing protein tau that is involved in Alzheimer’s disease. The protein spastin appears to be related to the abnormal placement of tau. This theory is that misplaced tau triggers an enzyme related to tagging with polyglutamine and this then causes the spastin to start breaking apart microtubules. Some findings related to kinesin motors are consistent with this as well.
Stabilization of Microtubule Structures
If a microtubule is cut, both pieces can start new lattices. Special proteins appear to notice the cut and find and cap the negative ends that stabilizer these new beginning structures. These caps stop further breakdown of the lattice. These proteins have been correlated with creating specific types of axons and dendrites and the production of asymmetrical neurons.
There are many different kinds of microtubule lattice structures that are quite different in terms of stability. Some have many proteins attached to hold the form, while others may use tubulin that has specific tags. One particular tag—polyamination—is critical for stability. Particular polyamines create special characteristics of the structure—putrescine, spermidine and spermine are examples. These very stable sections can be moved to other regions to create special very stable structures and compartments. There are other equally important mechanisms with other proteins, some connected to the Golgi.
Spatial Arrangements of Microtubule Structures
 From the centrosome, the plus ends grow out in a particular direction that leads into the axon initial segment and then the axon. It is not known how it understands to orient in straight parallel bundles. Those that are cut and stabilized are bundled with existing tubules. A particular kinesin motor has cross-link behavior and tau holds some bundles together. Those tubules that are not oriented properly are removed. Dynein motors help to emphasize the plus end version and push back the minus types. There are many different motors involved in complex ways. Kinesin-5 and 6 are motors that slide and coordinate the orientation of the tubules. These motors are critical in forming the mitotic spindle in the dividing cells, where microtubules pull chromatin in various ways. These functions occur with motors and with many specific associated proteins. They help with the opposite orientation in the dendrite, by sliding out the ones with the wrong orientation. Other motors are involved in creating branching of the lattice.
From the centrosome, the plus ends grow out in a particular direction that leads into the axon initial segment and then the axon. It is not known how it understands to orient in straight parallel bundles. Those that are cut and stabilized are bundled with existing tubules. A particular kinesin motor has cross-link behavior and tau holds some bundles together. Those tubules that are not oriented properly are removed. Dynein motors help to emphasize the plus end version and push back the minus types. There are many different motors involved in complex ways. Kinesin-5 and 6 are motors that slide and coordinate the orientation of the tubules. These motors are critical in forming the mitotic spindle in the dividing cells, where microtubules pull chromatin in various ways. These functions occur with motors and with many specific associated proteins. They help with the opposite orientation in the dendrite, by sliding out the ones with the wrong orientation. Other motors are involved in creating branching of the lattice.
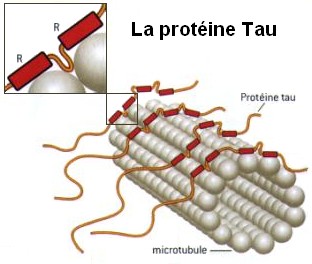
Most microtubules are in bundles, tied together by several molecules, most notably the well-known tau. Tau is in both axons and dendrites, whereas others are specific to one. Tau is particularly dense in the longer portions of the axon. It is tau that makes organization of the bundles appropriate and stable. Microtubule bundles grow straight in the dendrite but then have to dramatically turn to create spines. Actin with its special proteins and motors links the microtubules together at this branch points. Other mechanisms involve calcium signaling promoting actin structures that promote microtubule structures.
Are Microtubules the Brain of the Neuron?
The actions of microtubules are fantastically complex. They respond instantly in a vast amount of different ways to mental events in thousands of places throughout the neuron at once. How is this directed? How can anyone think that this is a random process? How can anyone not see that mental events direct these fantastically complex processes?