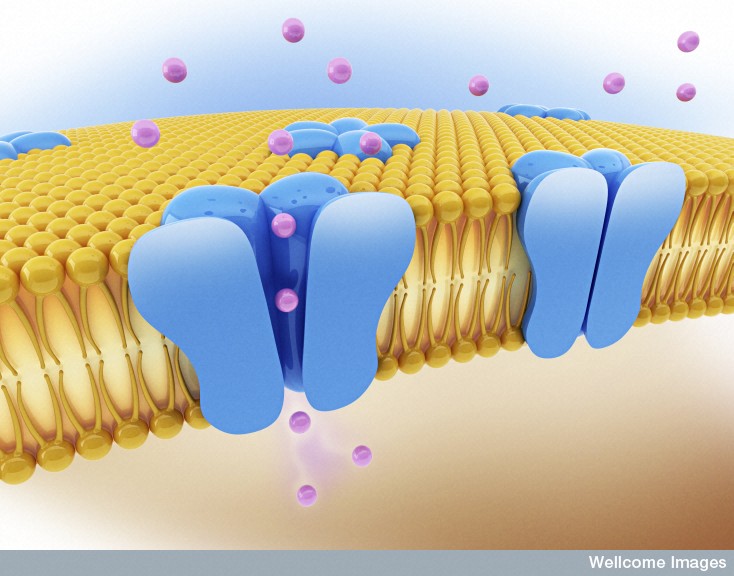
 Most students of biology are familiar with the neuron’s action spike traveling along the axon because of electric flux of the sodium and potassium channels in the membrane. It is not widely known, however, that there are almost a hundred different kinds potassium channels in the brain with very different properties. The many different potassium channels are constructed from a large number of subunits. They exist in varied compartments in the neuron. They are, also, different in the thousand different kinds of neurons in the brain (see post How Many Different Kinds of Neurons Are There). These potassium channels provide many varied functions and are found in multiple different compartments in proximal and distal dendrites and axons. Numerous types are specific to pre synaptic terminal regions, the cell bodies and at the nodes of Ranvier. With so many different types of neurons, it is only a little surprising that there would be a fantastic complexity in brain potassium channels
Most students of biology are familiar with the neuron’s action spike traveling along the axon because of electric flux of the sodium and potassium channels in the membrane. It is not widely known, however, that there are almost a hundred different kinds potassium channels in the brain with very different properties. The many different potassium channels are constructed from a large number of subunits. They exist in varied compartments in the neuron. They are, also, different in the thousand different kinds of neurons in the brain (see post How Many Different Kinds of Neurons Are There). These potassium channels provide many varied functions and are found in multiple different compartments in proximal and distal dendrites and axons. Numerous types are specific to pre synaptic terminal regions, the cell bodies and at the nodes of Ranvier. With so many different types of neurons, it is only a little surprising that there would be a fantastic complexity in brain potassium channels
 The neuron is unusual in that it uses two thirds of the genome to make a wide variety of proteins. It is, also, unusual in that one cell can be a yard long. The neuron’s complexity is based on many different unusual cellular compartments and the use of proteins to provide the unique constantly changing structures for neuroplasticity. Large families of proteins in the neuron include channel molecules in the membrane that allow control of the flux of different ions including sodium, potassium and calcium. These channel proteins are used as receptors for signals from other cells (inotropic receptors) as well as signaling inside the cell. Potassium channels are the third largest group of proteins used for signaling in cells. (GPCR and kinases are the other two).
The neuron is unusual in that it uses two thirds of the genome to make a wide variety of proteins. It is, also, unusual in that one cell can be a yard long. The neuron’s complexity is based on many different unusual cellular compartments and the use of proteins to provide the unique constantly changing structures for neuroplasticity. Large families of proteins in the neuron include channel molecules in the membrane that allow control of the flux of different ions including sodium, potassium and calcium. These channel proteins are used as receptors for signals from other cells (inotropic receptors) as well as signaling inside the cell. Potassium channels are the third largest group of proteins used for signaling in cells. (GPCR and kinases are the other two).
Almost A Hundred Different Potassium Channels
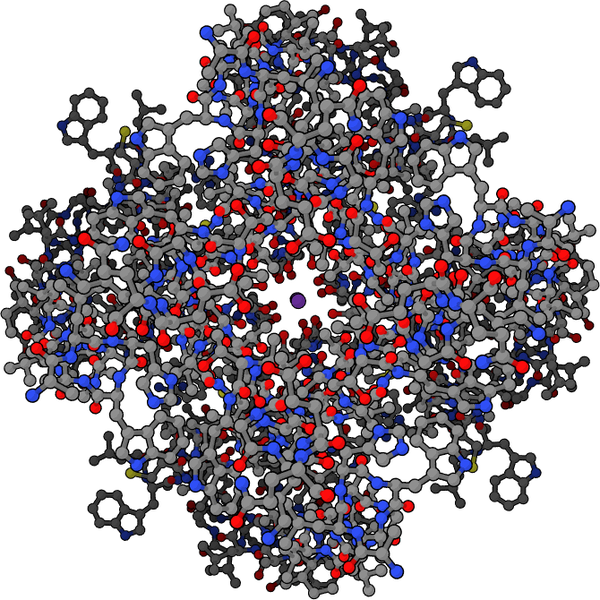 It is widely known that there are many different ionic receptors in synapses, such as sodium, calcium and inotropic glutamate receptors. But, it is not understood that there are, also, many unique potassium channels in each of the regions that are critical for neuronal transmission.
It is widely known that there are many different ionic receptors in synapses, such as sodium, calcium and inotropic glutamate receptors. But, it is not understood that there are, also, many unique potassium channels in each of the regions that are critical for neuronal transmission.
Eighty genes code for the basic channel subunits, called α units, with many additional proteins and cofactors forming a large potassium channel (KCh) complex. Various complexes with different subunits form unique types of channels throughout the brain. The exceptionally large number of different complex types is based on construction using either 2 or 4 different types of α units, along with multiple additional subunits.
Previous posts have described electrical phenomenon in and around neurons. They noted that the widespread conception of multiple action potentials being mathematically computed at the synapse is a simplification. Simple computation is upended by the vast complexity of different kinds of spikes, spatial and time factors, as well as many other synapse variables (see post The Limits of Current Neuroscience). In fact, there are a wide variety of local electrical gradients in neurons and along axons that have impact. The varied potassium channels have differing vital roles in these complex electrical events inside the neuron, along the axon and in between cells. The local potassium channel complexes respond to local environments and various other stimuli. Recently, several potassium channels have been shown to pick up both chemical and electrical signals serving as a coordinator of these different signals.
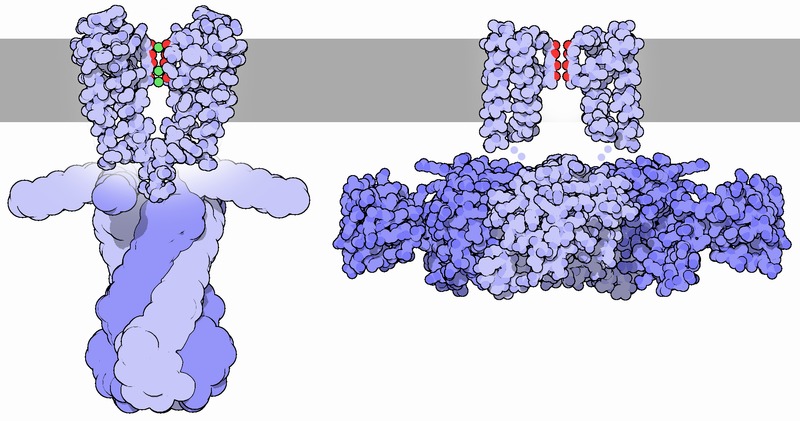
Potassium channels open and close allowing ions to move rapidly down a chemical gradient. They are specific to potassium and don’t allow sodium, calcium, magnesium or any other ion. They can produce the resting electrical potential for a cell and can reset the potential after an electrical event. The shape of the axon’s action potential is determined by the rate of the potassium flow after the sodium flow. In the heart, they keep the proper rhythm and when the general level of potassium is high or low, deadly heart rhythms can occur. They, also, maintain the tone of blood vessels and regulate secretion of insulin and other hormones. .
One overly simplified view of the types of channels includes at least 6 dendrite types, 2 axon and terminal types, 4 dendrite and soma types, 9 intracellular and 16 unknown. Each type has many variations.
A more comprehensive classification is available online as a resource ((https://www.ipmc.cnrs.fr/$duprat/ipmc/nomenclature). This site describes 15 superfamilies with more than 90 subtypes. The families are called KCNA, KCNB, KCNAB, KCNC, KCND, KCNE, KCNF, KCNG, KCNH, KCNJ, KCNK, KCNM, KCNN, KCNQ, and KCNS.
Subunits and Cofactors of Different Channels

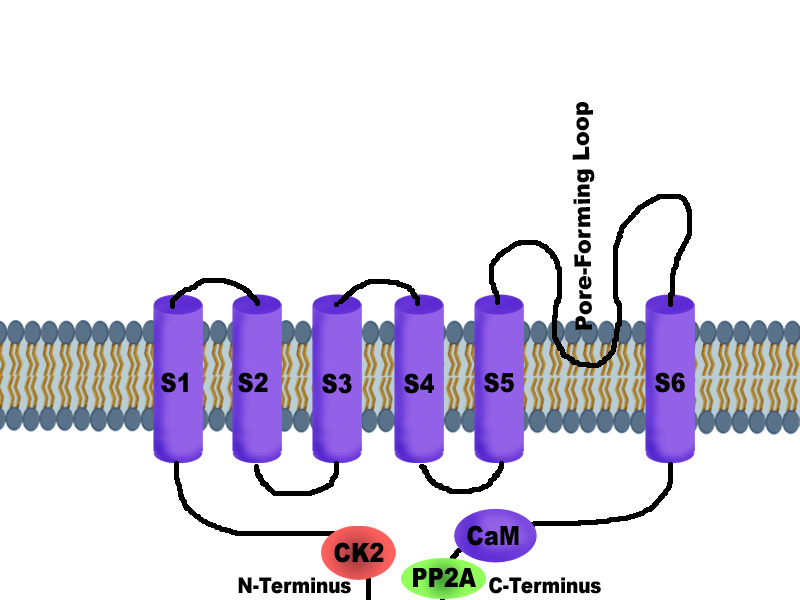
The primary subunits are called α units, which are proteins that cross the membrane multiple times (many cross six times, some four, two and one). They form a central pore that allows potassium to flow through or stops it from flowing. There are many other regions on the protein to provide many varied functions. The 80 genes manufacture 4 major types and many subtypes.
Four classes include:
- Channels triggered by calcium or other signals. These are called Kir and use G proteins and ATP in cascades (fifteen types).
- Channels that pass positive charge into the cell. These are called KCa and are activated by calcium and sodium (eight types).
- Channels that are always open or “leak” to provide a basal negative membrane potential. These are called K2P and are triggered by oxygen tension, pH, mechanical stretch, and G-proteins (fifteen types).
- Channels that are voltage gated responding to specific electrical gradients. These are called Kv (41 types).
Selectivity Filters
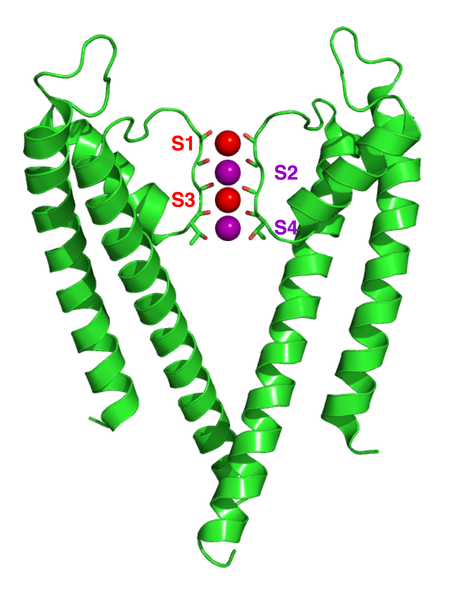 When potassium ions enter the channel they are in a complex surrounded by water molecules. The channel has a mechanism, called the selectivity filter, to remove the shell of water from the ion as it enters the P loop of the proteins that cross the membrane. All of the different types of channels have basically the same five amino acid sequences in the selectivity filter.
When potassium ions enter the channel they are in a complex surrounded by water molecules. The channel has a mechanism, called the selectivity filter, to remove the shell of water from the ion as it enters the P loop of the proteins that cross the membrane. All of the different types of channels have basically the same five amino acid sequences in the selectivity filter.
The unique structure of this P loop has special carbonyl oxygen molecules facing the center of the channel with a charge of negative electricity. The position of the multiple oxygen in this structure is similar to oxygen in water that normally surrounds the ion. This positioning creates a path for the ion to shed its water. The electrical activity of the rest of the loop draws the extracellular water solution into the channel to perform the function of freeing the potassium ion. In fact, it is a vastly complex mechanism that responds to many different signals such as calcium, electrical voltage and others.
Two alpha subunits combine to form some channels and there are four in most. With many subunits available, one group can assemble together and others cannot. Typically four subunits combine and then many co factors collect to make a complex. These complexes have different abilities, functions, modulations and places of operation.
 Manufacture of the subunits into a functioning channel is straightforward. But, it is not clear how different subunits and factors are present in particular regions of the membrane and how they are related to the pathways forming the specific types of membrane (see post on Amazing Complexity of Cellular Membranes).
Manufacture of the subunits into a functioning channel is straightforward. But, it is not clear how different subunits and factors are present in particular regions of the membrane and how they are related to the pathways forming the specific types of membrane (see post on Amazing Complexity of Cellular Membranes).
After creation of the protein subunits by messenger RNA, it is not at all clear how they can be positioned all along the very large axon membrane. It is not understood why some are produced and not utilized at particular sites within the neuron and what directs this decision. In the neuron, the axon membrane can extend for a yard. Somehow, all along the axon specific channels are placed in very discrete locations and amounts.
The reason little is known about the vast array of different potassium subunits is that they are difficult to study in the human brain. The way proteins are studied relies upon labeling of subunits and following them in their travels. Tagged subunits cannot be followed in the human brain as in animal models. But, recent data has been supplied using antibodies for particular subunits.
With the vast complexity of subtypes  and the trafficking involved, there is still little information available. It is possible that messengerRNA exists for these subunits along the axon. Ribosomes exist along the axon that manufacture the scaffolding microtubules.
and the trafficking involved, there is still little information available. It is possible that messengerRNA exists for these subunits along the axon. Ribosomes exist along the axon that manufacture the scaffolding microtubules.
What we currently know is based on the probes we have and the regions of brain that have been accessible to research; it is not based on the diversity and importance of the many types. A lot is known about the hippocampus and cerebellum and some particular unique types of neurons because the structures are simpler to observe. One easily studied neuron is called the Calyx of Held (looks like a flower calyx) in the auditory tract. Because of its size, potassium channels have been tagged by antibodies and seen under advanced microscopes.
Different Potassium Channels in Neuron Compartments
A previous post noted that there are probably thousands of different types of neurons. In addition, there are thousands more variations in the postsynaptic densities. So, it is not surprising that many different types of neurons utilize different types of channels placed in the unique anatomical regions associated with the dramatically different neuron structures. Differences in function have been observed, also, in the many different interneurons in different regions.
Potassium Channels in Dendrites
In the hippocampus CA1 region there are many different potassium channels that determine the specific electricity of the various dendrite arbors. The channels near the cell body and at a distance are quite different and have different levels of stimulation and excitation. Some form a gradient with varied channels in the dendrite branches. The channels near spines are unique. In fact, genetic alterations of these are related to neurological disorders. There are many variations related to proximity to the postsynaptic densities.
Similar types of channels are seen in areas outside of these hippocampal regions in pyramidal neurons and the dentate granule cells. These have different placements of specific subunits in areas that are away from the cell bodies and those that are close. One study showed that after a seizure, the locations of specific units are altered. This is possibly due to the unusual electrical activity produced, in part, by the channels. Or the effect of the abberant electricity on the subunits. Some of these changes appear to be protective in trying to reduce the chance of the unusual electrical activity and some appear to create seizures.
Distal Dendrites
Distal dendrites use specific channels to maintain low threshold for amplification of the signal coming from the axon.
Proximal Dendrites
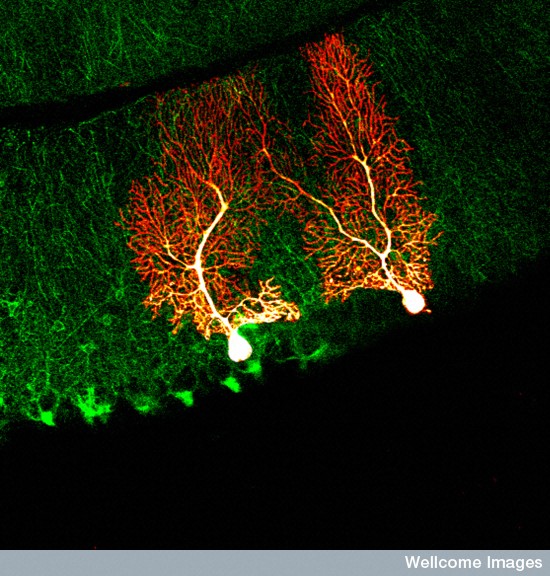 In proximal dendrites (closer to the cell body) specific amplification is determined by various different channel types. Also, unique channels provide support for the different types of vesicle transport at the synapse. These mechanisms use myosin II motors in the membranes (see post Yet Another Form of Neuroplasticity with Myosin Motors). The myosin II mechanism is critical to produce the particular channels specific for the proximal region. This effect is mediated by structures of actin scaffolding molecules that are connected with the myosin motors.
In proximal dendrites (closer to the cell body) specific amplification is determined by various different channel types. Also, unique channels provide support for the different types of vesicle transport at the synapse. These mechanisms use myosin II motors in the membranes (see post Yet Another Form of Neuroplasticity with Myosin Motors). The myosin II mechanism is critical to produce the particular channels specific for the proximal region. This effect is mediated by structures of actin scaffolding molecules that are connected with the myosin motors.
In the cell body itself there are a large amount of different channels that make its unique electrical activity for summation of the various inputs. It is thought that the calculation of the effects of various axon inputs is determined in this region. This region has unique plasma membrane structures with cisternae underneath. These are, also, connected with special calcium channels.
Axons and the Presynaptic Region
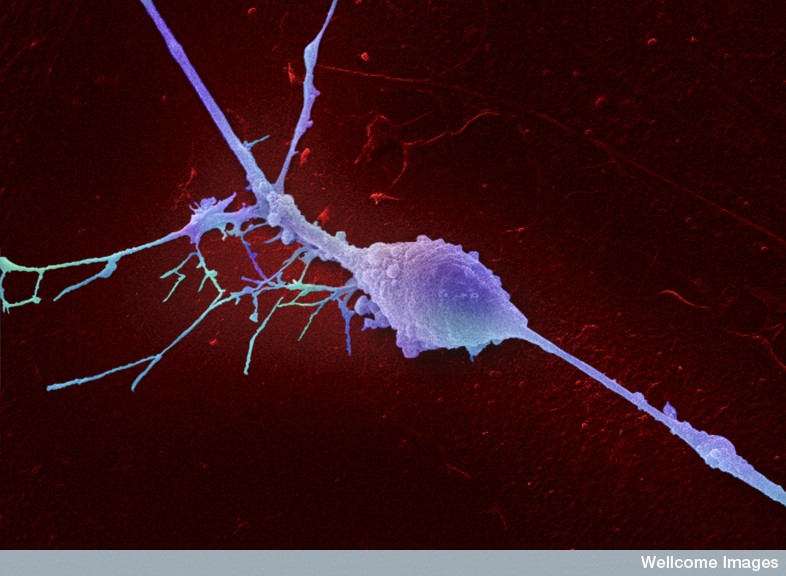 After the action potential, the potassium channels are critical to the way spikes approach the axon terminals. Some are involved in allowing the electrical activity to subside and some prepare for the next spike. For each of these functions there are different sets of potassium channels with varied subunits. Alterations in these particular channels cause many different diseases. New seizure medications are based on blocking some of the specific channels. Several unique channels are found in bipolar disorder.
After the action potential, the potassium channels are critical to the way spikes approach the axon terminals. Some are involved in allowing the electrical activity to subside and some prepare for the next spike. For each of these functions there are different sets of potassium channels with varied subunits. Alterations in these particular channels cause many different diseases. New seizure medications are based on blocking some of the specific channels. Several unique channels are found in bipolar disorder.
Beginning of the Axon
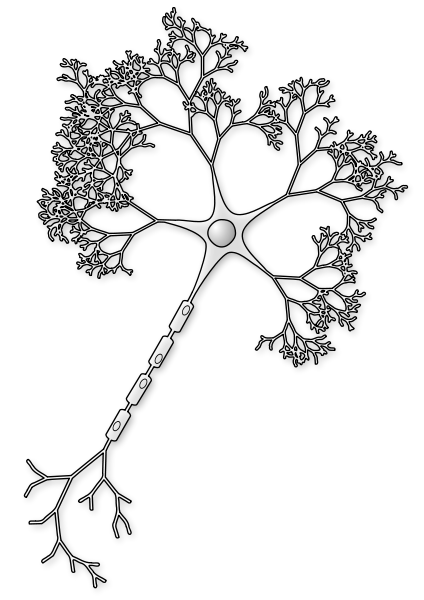 The first section of the axon is called the initial segment. Different potassium channels are critical to start or inhibit the action potential in this region. The large number of different subunits in this segment appears to be very significant for various neurological disorders such as genetic causes of epilepsy. This complex region is, also, changed in brain injury and disease causing yet other problems.
The first section of the axon is called the initial segment. Different potassium channels are critical to start or inhibit the action potential in this region. The large number of different subunits in this segment appears to be very significant for various neurological disorders such as genetic causes of epilepsy. This complex region is, also, changed in brain injury and disease causing yet other problems.
Further away from the initial segment there is yet another set of different channels. These determine the strength of the spike and are involved in repolarization (coming back to normal to prepare for another spike). Some of the channels are similar in both regions and many are uniquely different. There are specific channels that block the spike (retrograde) and that increase the spike (anterograde). Some are critical for the spike to be rapid onset and sharply defined.
Myelin and Channels
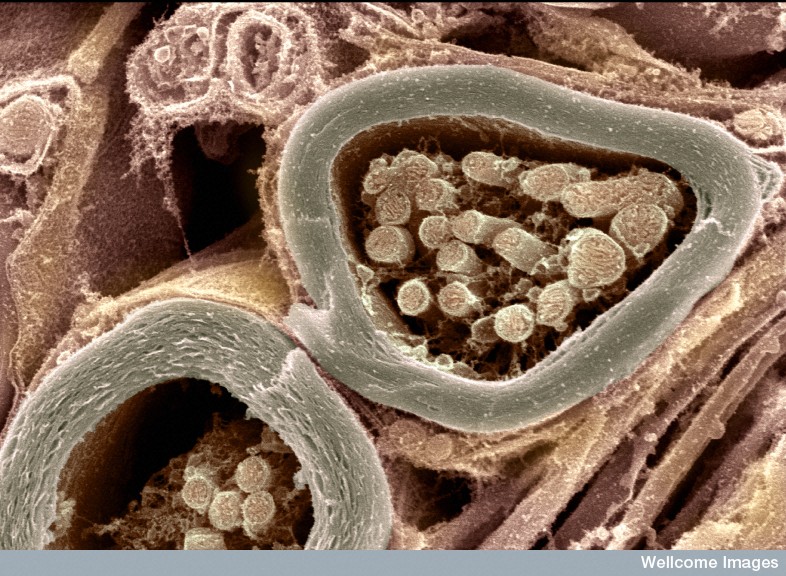 Specific channels reside near the nodes of Ranvier of myelinated neurons. The gap between the myelin at the internode is about 1 micrometer in size. At these nodes there is no myelin; the membrane participates in a unique type of electrical activity, called saltatory conduction, where the current jumps from node to node. For this activity there are many unique potassium channels in the internode and, also, just next to it (called the paranode) under the myelin sheath. There are no other potassium channels under the long stretches of myelin. The large amount of different potassium channels compared with sodium channels in the internode is quite different from the more equal proportion in the unmyelinated axons.
Specific channels reside near the nodes of Ranvier of myelinated neurons. The gap between the myelin at the internode is about 1 micrometer in size. At these nodes there is no myelin; the membrane participates in a unique type of electrical activity, called saltatory conduction, where the current jumps from node to node. For this activity there are many unique potassium channels in the internode and, also, just next to it (called the paranode) under the myelin sheath. There are no other potassium channels under the long stretches of myelin. The large amount of different potassium channels compared with sodium channels in the internode is quite different from the more equal proportion in the unmyelinated axons.
Until recently, research only focused on the sodium channels and it was not clear that potassium channels have important functions in saltatory conduction. Potassium channels now appear to be critical and regulate the rapid repolarization of the action potentials.
There are many different potassium channels in the internode and just near the node. The channels under the myelin near the edge of the internode prevent firing and help the internode’s resting potential to be prepared for the saltatory spike. A previous post described the new complexity found in the myelin code. It showed that only specific sections of cortical axons are myelinated. Now, there is even greater complexity with many different potassium channels.
Pre synaptic Terminals
 Unique potassium channels are involved in regulating the release of the vesicles with neurotransmitters. One group is at the region just before the terminal section. These are part of filters or gatekeepers for the approaching electric spike. They can inhibit the effect of a spike. Once in the presynaptic terminal, channels with different subunits regulate the level of electrical activity and the release of vesicles with neurotransmitters.
Unique potassium channels are involved in regulating the release of the vesicles with neurotransmitters. One group is at the region just before the terminal section. These are part of filters or gatekeepers for the approaching electric spike. They can inhibit the effect of a spike. Once in the presynaptic terminal, channels with different subunits regulate the level of electrical activity and the release of vesicles with neurotransmitters.
Other specific channels repolarize in the terminal. A very unique new form of neuroplasticity has been described that involves specific potassium channels and a derivative of arachidonic acid (see posts on cannabinoids and fatty acids) causing retrograde messengers.
Coupling Diverse Channels
 Not only are the many potassium channels coordinated amongst themselves, but, also, with other types of ion channels, such as sodium and calcium.
Not only are the many potassium channels coordinated amongst themselves, but, also, with other types of ion channels, such as sodium and calcium.
Neurons have 10,000 times greater membrane area than other human cells. But, even with this huge membrane, the localization of the potassium channels is extremely precise and very specific. Each tiny discrete location uses unique types of potassium channels. One example is an axon that goes from the cortex to the lower spine that can be a yard long. A particular type of critical potassium channel exists in a region that consists of 0.1% of the membrane. In the dendrites, the regions are even smaller. These specific channels are brought to these places and then maintained at specific levels. It is not at all clear how this is all directed. Most of the 90 channels are yet to be clearly defined.
Many unique patterns of potassium channels appear in specific neuron regions. There is a high density of clusters of both sodium and potassium channels in the myelin internode. There are high densities in the pre terminal regions. In terminal regions, there are a few sodium channels and more potassium versions. Other unique patterns are just now being discovered. Many patterns form unique associations with calcium channels using particular types of potassium subunits in particular spatial arrangements.
Fantastic Complexity in Brain Potassium Channels
 Of the recently discovered 90 different potassium channels, there is great specificity of their placement in exact patterns and in very distinct regions of the neuron. Some channels exist along the entire axon; most do not. Very specific channels are placed at the cell body, the beginning of the axon, further along the axon, at myelin internodes, at distal dendrites, at proximal dendrites, near the post synaptic density, near the calcium channels at the synapse and near the sites of neurotransmitter vesicle release. They are in unique patterns for many of the 1000 different kinds of neurons.
Of the recently discovered 90 different potassium channels, there is great specificity of their placement in exact patterns and in very distinct regions of the neuron. Some channels exist along the entire axon; most do not. Very specific channels are placed at the cell body, the beginning of the axon, further along the axon, at myelin internodes, at distal dendrites, at proximal dendrites, near the post synaptic density, near the calcium channels at the synapse and near the sites of neurotransmitter vesicle release. They are in unique patterns for many of the 1000 different kinds of neurons.
Phosphorylation and kinase cascades are involved in producing different pathways for subunits. Some are produced by specific interactions of multiple proteins such as binding of calmodulin and calcium effects. But, by and large the phenomenal amount of differences and movement of these varied subunits are regulated in an unknown manner. Axons can be a yard long or tiny. In either case, very specific potassium channels are placed in the membrane at very exact places.
With this fantastic new complexity of potassium channels, the question again must be asked where the direction for all of this resides? The activity in all of the neuron’s potassium channels is triggered by, and is related to, mental events. Is there a mental connection to these specific molecules?