 White blood cells, which are called leukocytes, are able to find sites of infection and trauma through elaborate signaling. Of the many different kinds of immune and blood cells, the particular type is produced and sent long distances to the site of the trouble. First, they travel in blood vessels and then into the tissue, despite many obstacles. New information shows that leukocyte travel and the signaling producing it is even more complex than reported in a previous post. Leukocytes are now seen to travel backwards away from the inflammation or injury after the job is done. All the dramatic types of travel are completely controlled by signals from many participating cells in the blood vessels, the tissue, and the immune and nervous systems.
White blood cells, which are called leukocytes, are able to find sites of infection and trauma through elaborate signaling. Of the many different kinds of immune and blood cells, the particular type is produced and sent long distances to the site of the trouble. First, they travel in blood vessels and then into the tissue, despite many obstacles. New information shows that leukocyte travel and the signaling producing it is even more complex than reported in a previous post. Leukocytes are now seen to travel backwards away from the inflammation or injury after the job is done. All the dramatic types of travel are completely controlled by signals from many participating cells in the blood vessels, the tissue, and the immune and nervous systems.
Recently, new devices have allowed better observation of this complex travel through infected and injured organs. Previously, these cells were considered to be short lived thought to commit programmed suicide after their tasks. Now, more complex behavior has been observed. Leukocytes send very elaborate signals from the site and even return to the blood for more work in other places.
Migration toward and away from sites of inflammation are determined by a variety of complex signals. The previous post demonstrated many different ways that these cells are able to travel over difficult terrain, sometimes even without oxygen. This post will update this information with even more complex signaling. First, there is a brief summary of the previous post on leukocyte methods of travel.
Summary From Previous Post on Leukocyte Travel
 The previous post elaborated on exact methods of travel. When cells move through the body, complex signals accompany them from lining endothelial cells, blood vessel cells, immune cells, and neurons. Leukocytes and T cells are able to use a wide variety of different methods to travel to critical sites of injury and infection and then enter into the specific organs of the human body.
The previous post elaborated on exact methods of travel. When cells move through the body, complex signals accompany them from lining endothelial cells, blood vessel cells, immune cells, and neurons. Leukocytes and T cells are able to use a wide variety of different methods to travel to critical sites of injury and infection and then enter into the specific organs of the human body.
Techniques include changing shapes and using the leading edges of the cell in different ways. Leukocytes can decide to suddenly swarm near a target, like insects. They use chemical gradients, cytokine signaling with cells and extracellular matrix. Cells are able to alter their shape and modes of transport as needed. Leukocytes use back and forth communication with many other cells to analyze the best way to traverse specific terrain, as well as the specific response to the target problem.
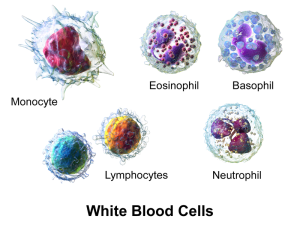
Leukocytes use very complex modes of travel to navigate the vastly different environments of the various human organs. Neutrophils are a variety of leukocytes that can eat debris and travel throughout the body, in and out of these tissues. They grab onto endothelial lining cells of blood vessels with adhesion molecules that allow rolling, tethering, and firm adherence.
 Leukocytes must travel between lining cells and through the dense basement membrane into the extracellular space where they meet very different factors including chemical gradients that attract and repel them. To transit the basement membrane, signals from other cells are required to alter membranes. Leukocytes alter shape as well as direction many times. Actin-based propulsion allows travel along non-cellular scaffolding structures. Once at the infected or damaged sites, they take stock and provide many repair functions.
Leukocytes must travel between lining cells and through the dense basement membrane into the extracellular space where they meet very different factors including chemical gradients that attract and repel them. To transit the basement membrane, signals from other cells are required to alter membranes. Leukocytes alter shape as well as direction many times. Actin-based propulsion allows travel along non-cellular scaffolding structures. Once at the infected or damaged sites, they take stock and provide many repair functions.
Individual cells have multiple different kinds of movement for different three-dimensional environments, provided by operation of specific actin-myosin motors, scaffolding structures, and transit tubules.
- Amoeboid movement uses a round cell shape and a leading edge that moves. The pseudopod is a gliding form of movement, maintaining a basic round shape, especially in the rear. Actin builds large molecules that rapidly move the leading edge forward into arms called pseudopods. Then other myosin motors contract in the middle of the cell. A large amount of different myosin motors in the back do not allow protrusion, which pushes movement forward.
- Blebbing movement is another form that has contractions at the rear of the cell. This pushes cytoplasm forward forming round out-pouchings called blebs. This type of movement depends on outside adhesion.
- Adhesions from traveling cells attach to the lattice of the extracellular matrix allowing the cell to protrude forward. The attachment is by integrin molecules and a linking mechanism called the molecular clutch, which engages and disengages rapidly. This apparatus uses actin moving backwards and linking to integrin while the extra cellular matrix pushes the cell forward.
 Recent research shows that leukocytes walk stepwise with two feet below that alternatively form adhesions in different places. When the scaffolding proteins reach forward, these adhesions help push the cell.
Recent research shows that leukocytes walk stepwise with two feet below that alternatively form adhesions in different places. When the scaffolding proteins reach forward, these adhesions help push the cell.- Leukocytes (dendritic cells, neutrophils, and lymphocytes) use an actin protruding, integrin linking adhesion mode of movement some of the time and an independent form without attachment to extracellular structures other times. They can move along adhesive and non-adhesive surfaces at similar speeds.
- When the tissue is very dense they must make special actin-myosin contractions in the rear. Leukocytes are able to rapidly change their shape for different situations and travel quickly through many different environments.
- In low-density matrices, they can use less myosin activity. In highly dense material they form another structure called pearl-chain nuclei. Here they change the shape of the nucleus to get through small crevices rather than the typical large oval nucleus. Very recently, research has shown that leukocytes can move using scaffolding either when stimulated by external signals, or spontaneously with out external direction. In this latter case, the cell determines a specific direction to move.
 Recent research shows that leukocytes walk stepwise with two feet below that alternatively form adhesions in different places. When the scaffolding proteins reach forward, these adhesions help push the cell.
Recent research shows that leukocytes walk stepwise with two feet below that alternatively form adhesions in different places. When the scaffolding proteins reach forward, these adhesions help push the cell.Neutrophils Are the Most Common White Blood Cells
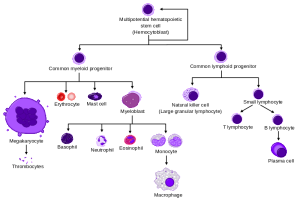
Neutrophils are the most common white cells in the blood and they are prominent as the first responders to trauma and infection. Neutrophils use a wide variety of techniques to manage the infection. They are able to eat debris and microbes through phagocytosis.
Neutrophils use toxic chemicals that are stored in their prominent granules. They produce molecules called reactive oxygen species (ROS). Oxygen is extremely reactive, even dangerous in the body, and has to be carefully monitored. Producing reactive oxygen molecules is used to increase inflammation response and to kill microbes. ROS are also used as regulatory signals.
Neutrophils can build unusual structures called neutrophil extracellular traps (NETs) in the extracellular space. They consist of fibers made of DNA, proteins, and other material from neutrophil granules. They become a net or mesh to capture microbes where the toxins kill them.
When there aren’t enough leukocytes, infections are severe and recurrent. But, too many leukocytes also cause problems such as too much inflammation with ensuing tissue damage.
Neutrophils use many signals and produce many different kinds of enzymes that cut proteins, called proteases. They send signals to T cells and B cells to get them involved, but there is a fine line between enough and too much. Among other regulatory immune cells, neutrophils are now known to be key players in regulating these very complex processes.
Neutrophils go through one phase where they are first called into duty. In this phase they send many different signals. Then in another phase, they must focus all the issues involved in resolving the particular type of inflammation and trauma. This phase can include neutrophils reversing directions and leaving the area.
Moving Forward
 When damage occurs, a variety of cells start signaling. A previous post showed that even platelets (who are the real first responders to damage) engage in elaborate signaling. They set up gradients of molecules that become directional signals. Leukocytes must make sense of the chaos that damage produces.
When damage occurs, a variety of cells start signaling. A previous post showed that even platelets (who are the real first responders to damage) engage in elaborate signaling. They set up gradients of molecules that become directional signals. Leukocytes must make sense of the chaos that damage produces.
The first phase of response to the damage is called scouting. After this, signals are increased in an amplification phase. These signals are from both blood and immune cells and local tissue cells as well. This second phase brings in more cells from blood vessel using receptors for damage-associate molecular patterns or DAMPs, hydrogen peroxide, metabolites of fat tissues, and special chemokine signals giving directions.
Migration is not simple and many different signals are involved. No one signal is sufficient for migrating. Neutrophils have more than 30 different receptors related to sensing inflammation, directional signals, and altering behavior. Some of these receptors are G protein coupled receptors (GPCRs). These also include adhesion receptors that guide leukocytes along certain paths. They include many cytokines that stimulate other networks of cells and pattern recognition receptors (PRRs). Signals alter behavior related to eating debris, using granule molecules and producing ROS. They also can signal cell suicide. Many different pathways are triggered inside the cell.
Early Signals
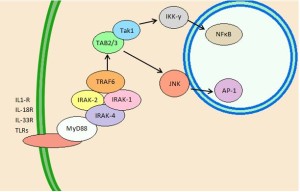
Early signals are from damaged tissue cells, most often DAMP receptors. Leukocytes pick up signals from recognition receptors such as TLRs (Toll like receptors) and NOD like receptors (NLRs). What they pick up are pieces of DNA, proteins, ATP and other signals.
One of the first signals is H2O2, which stimulates leukocytes to move, triggering many pathways. It lasts less than an hour. Longer signals are needed to sustain stimulation.
Damaged cells release many molecules (DAMPs) including ATP, which are picked up by many receptors. They affects mitochondria to produce more energy. Damaged mitochondria release peptides that are signals as well. These peptides become a strong local signal for immediate localization.
A large family of chemokine signals is vital for leukocyte migration (CSCL1, 2, 3, 5, 6, 7, and 8). T cells, macrophages and neutrophils, produce these chemokines. CXCL8 attaches to cell walls and extracellular matrix to form gradients of directions for travel.
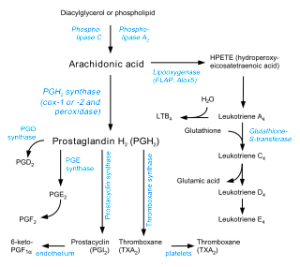
Fat molecules metabolized from arachidonic acid are significant signals as well. A previous post on cannabis and omega 3 and 6 described the large number of metabolites that are lipid based signals. Of all of these lipid signals, leukotriene B4 known as LTB4 is the most powerful to attract leukocytes. It starts early in the process and is used to greatly enlarge the response as well as in the next phase. Leukotrienes are the type of cytokines that work with leukocytes.
Many lining cells maintain large amounts of stores of these signaling molecules just in case. DAMPs stimulate rapid release after trauma and cell damage. This damage also releases ions from the cells altering the environment dramatically.
Amplifying Responses
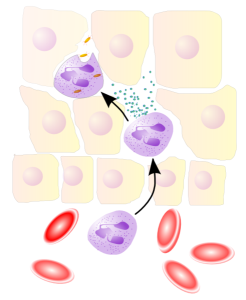
One aspect of amplification is “neutrophil swarming. After traveling from the blood vessel into tissues, neutrophils exhibit very complex coordinated clustering formations that have been likened to insect swarming behavior and hence the term neutrophil swarming. The early responding neutrophils and macrophages send large amounts of signals that also increase the signaling of the tissue cells. These signals promote movement of leukocytes forward toward the trouble. Cells create small gradients of the LTP4 that greatly enhances other signals. This also keeps the directions of travel aligned. Much bigger gradients can also occur. LTB4 seems to be critical for signaling of an increasing number of leukocytes and all of them join in this production. The original peptides also stimulate more LTB4 in this phase.
Some of the original cells die as part of this process of greatly enlarging the swarm. Producing dead cells is a technique of creating more damage molecules (DAMPs) to continue the enlargement of the stimulation cycle for more leukocytes. This cycle continues where dead cells produce DAMP particles that stimulate even more cytokines and leukocytes.
Similar amplification events occur in arthritis with the same cytokines that stimulate interleukin (IL-1β). This cytokine greatly increases stimulation of the vessel lining and tissue cells, fibroblasts and other immune cells such as macrophages.
Neutrophils produce enzymes (metalloproteinases), which cut the CXCL cytokine molecules producing even greater effects. These enzymes also cut collagen extracellular molecules that become signals to enlarge cells being attracted. It appears that all types of immune cells join in this signaling.
Different Signals for Trauma and Infection
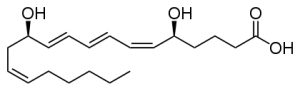 There are differences between responses from inflammation and traumatic injury. Inflammation is stimulated until the infection is gone. But, white blood cells must be controlled in infection because they can produce microenvironments where microbes can breed. Microbes also can alter the neutrophil responses and kill them producing signals for increased inflammation. Pieces of material from microbes are called pathogen-associated molecular patterns (PAMPs) and this creates another round of signaling. These signals call for a much wider range of immune cells and much more complex response, triggering T cells, macrophages, dendritic cells and mast cells.
There are differences between responses from inflammation and traumatic injury. Inflammation is stimulated until the infection is gone. But, white blood cells must be controlled in infection because they can produce microenvironments where microbes can breed. Microbes also can alter the neutrophil responses and kill them producing signals for increased inflammation. Pieces of material from microbes are called pathogen-associated molecular patterns (PAMPs) and this creates another round of signaling. These signals call for a much wider range of immune cells and much more complex response, triggering T cells, macrophages, dendritic cells and mast cells.
When trouble starts as trauma and then infection starts, more signals call for neutrophils. In this situation, all immune cells signal for neutrophils. Macrophages signal for entry from blood vessels and mast cells call them directly to the site. Back and forth communication of immune cells, tissue cells and leukocytes tackle the infection.
 Mast cells also use multiple leukotrienes including LTB4 and LTC4. Multiple immune cells form a feed forward feedback loop all greatly increasing the volume of signals and attracting leukocytes from all over. There are cascades where a first responder calls for some, then macrophages, and many other immune cells join in.
Mast cells also use multiple leukotrienes including LTB4 and LTC4. Multiple immune cells form a feed forward feedback loop all greatly increasing the volume of signals and attracting leukocytes from all over. There are cascades where a first responder calls for some, then macrophages, and many other immune cells join in.
T cells are vital in specific infections, such as strep pneumonia and H. pylori. They signal IL-17. This cytokine is critical to call more leukocytes by stimulating all tissue cells to signal for more with many different cytokines. Some of the signals stimulate bone marrow to make many more cells.
 Infections need much larger amounts of white cells. For clean wounds, too many cells are trouble. The ways that the tissues know whether it is one or the other is quite complex and just being discovered. Cells know to respond differently to DAMPs (clean wound) than PAMPs (with microbes). H2O2 and IL-1β are particular signals with clean wounds.
Infections need much larger amounts of white cells. For clean wounds, too many cells are trouble. The ways that the tissues know whether it is one or the other is quite complex and just being discovered. Cells know to respond differently to DAMPs (clean wound) than PAMPs (with microbes). H2O2 and IL-1β are particular signals with clean wounds.
Leukocytes travel differently to different signals. CD24 signals have multiple effects where they don’t allow DAMPs to hit the receptor. Through complex pathways it decreases NF-kβ, which would activate dendritic cells. In this way it decreases the calls for leukocytes. The types of neutrophils that come to wounds and infections are different as well. It is not clear if those that arrive are altered at the site, but they are distinct and respond differently.
Leukocytes Travelling Backwards
 One of the new unusual findings is that when injuries are repaired, a decrease of inflammation is essential. Leukocytes are removed from the site in severa newly discovered ways. Previously it was known this can occur by cell death either programmed suicide or killed by other cells. Macrophages can eat this debris.
One of the new unusual findings is that when injuries are repaired, a decrease of inflammation is essential. Leukocytes are removed from the site in severa newly discovered ways. Previously it was known this can occur by cell death either programmed suicide or killed by other cells. Macrophages can eat this debris.
Now, it has been found through enhanced observation techniques, that neutrophils reverse direction and travel out of the inflammation region once it is under control. This reverse travel takes them back into the blood. This new process is called rTEM or reverse trans-endothelial migration. These cells can then travel quite far away from the site.
Finding the signals that cause neutrophils to leave the scene of the injury is a new focus for medications. These cells re locate at various other places in the body and at times start or add to inflammation at these other sites. This movement appears to be more than a simple reverse attraction from a chemical gradient. Signals from inside the wound and from the blood vessels are part of the process. Neutrophils are able to take in a wide variety of signals and determine that it is time to leave the site of the injury.
While CXCL8 is one of the major attracting cytokines, when the concentration levels become great, the signal can be reinterpreted in reverse. Receptors in neutrophils become less activated by internal pathways. Another way that they are stimulated to leave is by direct contact with local macrophages. Macrophages appear to signal the need for leukocytes to tone down their work. It appears that signals from the macrophages affect both leukocytes and the macrophages themselves. Abnormal signals of this type might be relevant in excessive neutrophil inflammation in granulomas.
 Another signal for reverse travel is when the lipid metabolite and signal LTB4 changes to a different lipid based signal called LXA3 (lipoxin A4). These signals move the leukocyte in the opposite direction. They stimulate a variety of other lipid signals that also help leukocytes to move away from the site.
Another signal for reverse travel is when the lipid metabolite and signal LTB4 changes to a different lipid based signal called LXA3 (lipoxin A4). These signals move the leukocyte in the opposite direction. They stimulate a variety of other lipid signals that also help leukocytes to move away from the site.
Leukocytes can move away through the tissue or enter into the blood stream with the rTEM process through complex pathways. Multiple different mechanisms contribute to the decrease at the site.
Neutrophils in the tissue are noted to be using different genetic networks than those in the blood stream. Those that have used rTEM are much less likely to kill themselves and they produce more oxygen species products. Another group that travelled far in blood had different characteristics. Future research will need to identify these characteristics because these blood cells can contribute to inflammation elsewhere.
Recently in a study of pancreatitis, the lung had increased inflammation from these cells. Some have travelled through lymph into nodes and bone marrow stimulating these tissues. These cells can also carry microbes to these sites triggering T and B lymphocyte activity.
Targeting Migration for Treatments
 These traveling leukocytes can contribute to arthritis, pulmonary fibrosis and other inflammatory diseases. Medical treatment tries to resolve inflammation at a site without suppressing immune function that can be dangerous elsewhere. Too many dead cells can create their own problems. If new treatments can stimulate forward movement and reverse migration some of these problems can be dealt with.
These traveling leukocytes can contribute to arthritis, pulmonary fibrosis and other inflammatory diseases. Medical treatment tries to resolve inflammation at a site without suppressing immune function that can be dangerous elsewhere. Too many dead cells can create their own problems. If new treatments can stimulate forward movement and reverse migration some of these problems can be dealt with.
One treatment involves cholchicine, which inhibits microtubule function, stops forward movement of neutrophils and keeps neutrophils from accumulating at sites. Stopping the early phase of attraction signals might also be helpful. Stopping some of the amplification could be helpful using CXCR1 and CXCR2 inhibitors with cancer and autoimmune disease.
Other new drugs are targeting reverse migration as well. One new drug (tanshinome IIA) is from a Chinese herb. This herb increases suicide of neutrophils and causes more reverse migration.
Signals for Complex Travel of White Blood Cells
 Leukocytes and T cells use many techniques to travel in blood vessels and then into tissues to sites of infection and trauma. They are able to traverse unusual terrain sometimes without oxygen and with dense extracellular matrix. Signals are necessary at every step of the way. Signals first call for the type of cells needed from bone marrow. Signals direct them to the organ via blood vessels. Signals tell the lining cells and basement membrane to allow them into the tissues. And it is signals that direct them to fight the microbes or rebuild the tissues.
Leukocytes and T cells use many techniques to travel in blood vessels and then into tissues to sites of infection and trauma. They are able to traverse unusual terrain sometimes without oxygen and with dense extracellular matrix. Signals are necessary at every step of the way. Signals first call for the type of cells needed from bone marrow. Signals direct them to the organ via blood vessels. Signals tell the lining cells and basement membrane to allow them into the tissues. And it is signals that direct them to fight the microbes or rebuild the tissues.
Recently, with new tools to observe cell movement, it has been discovered that when the job is done, leukocytes are directed to move backwards out of the inflammation site. This new signaling is the focus of intense research for possible new medical treatments.