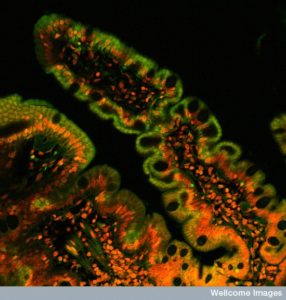
 With trillions of microbes comprising thousands of species or more, it has been impossible to fully study their composition or exact locations in the gut. Even more complex is their back and forth chatter between multiple Kingdoms such as bacteria, viruses, archaea, fungi, and eukaryote human cells in the lining and immune system. This conversation determines the outcomes of infections and their friendly stable life long communities. Very recent studies show that along the extensive human gut, there are thousands of different environments and niches, with different relationships between lining and immune cells and various microbe communities that are allowed to live nearby and help protect against dangerous infections.
With trillions of microbes comprising thousands of species or more, it has been impossible to fully study their composition or exact locations in the gut. Even more complex is their back and forth chatter between multiple Kingdoms such as bacteria, viruses, archaea, fungi, and eukaryote human cells in the lining and immune system. This conversation determines the outcomes of infections and their friendly stable life long communities. Very recent studies show that along the extensive human gut, there are thousands of different environments and niches, with different relationships between lining and immune cells and various microbe communities that are allowed to live nearby and help protect against dangerous infections.
These studies demonstrate a very complex situation where there are different kinds of settlements at each stage along the stomach, small intestine, cecum, appendix and large intestine. These three-dimensional villages, patches and cities are different in the moving center of the lumen, close to the mucus layer and along side the epithelial lining and its deep crypts. Biofilm communities exist in some regions and are not, in fact, single species either. Biofilms are complex multi faceted combinations of species (to be discussed in the next post).
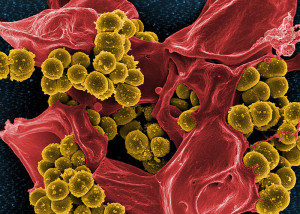
Studying fecal samples (the clinical gold standard to tell what species are present) tells very little because they destroy the three dimensional structures and critical spatial relationships between patches of different species. Painstaking research using biopsies all along the lining and extremely difficult studies that preserve the three-dimensional colony structures in stool samples are beginning to elucidate the vast civilizations and extremely complex communication in the microbe geo politics of the gut.
Recent research shows that if communities are too cooperative with each other they create an unrealistic growth pattern that fails. There has to be a balance of some cooperation and some competition for the communities to be stable for the long term. This post discusses research findings thus far about the geography of microbes along the gut.
Different Environments at Each Stage
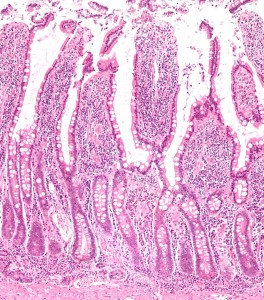 Various strains can survive based on very different environments. A further distribution occurs in each area for those in the middle of the lumen, in the mucus protecting the intestinal epithelial cells and in special indentations called crypts. Many factors influence which communities thrive in these ecological niches, including the level of acid and base, types of food that are present, and communication with different epithelial cells (see post Intelligent Intestinal Epithelial Cell) and various immune cells.
Various strains can survive based on very different environments. A further distribution occurs in each area for those in the middle of the lumen, in the mucus protecting the intestinal epithelial cells and in special indentations called crypts. Many factors influence which communities thrive in these ecological niches, including the level of acid and base, types of food that are present, and communication with different epithelial cells (see post Intelligent Intestinal Epithelial Cell) and various immune cells.
The infant microbe communities start with contact in the mother’s vagina and breast milk. They develop based on exactly when the baby is born during its fetal development. The milk gives the infant particular antibodies as well as other molecules, such as oligosaccharides, which provide food for particular microbes. The development of permanent colonies is based on the particular immunity, diet, exposures to medications and chemicals and the original vaginal microbes. This is all complicated by the fact that microbes provide food for each other, often established by the first group. The immune system secretes unique molecules that help certain microbe groups, such as immunoglobulin A. A very powerful factor is diet and the availability of specific food and its metabolites in different regions.
 Some microbes prefer specific oligosaccharides. Milk is influential in determining which colonies survive. When solid food begins, adult microbes grow. They consist of up to thousands of different types of microbe colonies. Many of these species are not found anywhere else. These have been divided into classes based on how they relate to the human host. Symbionts have a very close interaction but can include parasites. Commensals have a symbiotic relationship but only one-sided benefits. Relations with mutualists benefit both.
Some microbes prefer specific oligosaccharides. Milk is influential in determining which colonies survive. When solid food begins, adult microbes grow. They consist of up to thousands of different types of microbe colonies. Many of these species are not found anywhere else. These have been divided into classes based on how they relate to the human host. Symbionts have a very close interaction but can include parasites. Commensals have a symbiotic relationship but only one-sided benefits. Relations with mutualists benefit both.
Recent studies show that when a mouse gut is completely cleaned of all microbes and then particular microbes are added, these do not determine the communities that will develop later. Rather the permanent communities develop from particular factors that are present in the host and interactions with other communities. The human gut microbes are somewhat constant with approximately 40 to 50 prominent species making up three quarters of the volume. Another study found that species were basically stable for five years. When antibiotics are given the variety is limited for a time and then they return as before. Even if some species are not measurable, they return probably from special hiding places or niches.
Special Regions Are Sheltered

Three regions are somewhat isolated from the general communities in the intestinal stream—the layer of mucus that is near the epithelial cells, deep crypts or invaginations in the intestinal lining and the appendix.
Many different independent regions exist along the small and large intestine (colon). A special region connects the small and large intestines called the cecum, with a special valve, the ileocecal valve (ICV). The cecum is a pouch at the beginning of the large intestine and is a special environment for bacteria. In some animals, bacteria break down cellulose in this pouch, but not in humans. The appendix is even more protected and is attached to the cecum.
The small intestine has an acid environment from the influx of bile acids. It, also, has more oxygen and more antimicrobial molecules than other regions. All of these can kill microbes. The special microbes that adapt to this environment grow rapidly and can use oxygen. They are eager to have the carbohydrates that are available from breakdown of food. Bile acids have a dramatic effect on microbes in the small intestine and act as detergents decreasing surface tension. Because of the rapid liquid flow in the center of the small intestine (much less than the colon), microbes that can be part of the mucus layer (cells that line the mucus layer are called mucosa) or attach to the lining are much more likely to be able to live there for a long time.
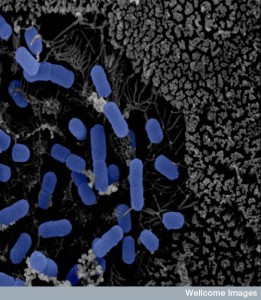 Microbes in the ileus have a large number of special genes related to human metabolism, such as dealing with sugars. There are more Proteobacteria, lactobacillae and Clostridium spp. These bacteria compete for the sugars but are limited by the bile acids. Near the end of the ileum there are more and more bacteria to the point of saturation, such as occurs in the large intestine.
Microbes in the ileus have a large number of special genes related to human metabolism, such as dealing with sugars. There are more Proteobacteria, lactobacillae and Clostridium spp. These bacteria compete for the sugars but are limited by the bile acids. Near the end of the ileum there are more and more bacteria to the point of saturation, such as occurs in the large intestine.
The slow moving cecum and colon have the greatest amount of different bacteria species. For a long time, the thin tube like appendix was considered useless. This was an opinion of those who believe that all evolution is completely random, since they couldn’t decipher a purpose for it. In fact, it is a unique niche that houses the greatest diversity of microbes in a very protected environment. The appendix is off of the main flow and doesn’t succumb to disease and antibiotics as much as the rest of the intestine. It, therefore, can be a resource to repopulate the large and small intestines after problems that occur through disease.
 The cecum and large intestine are able to break down and digest plant fibers. Recently, it was found that eating fiber early in life is necessary to maintain diverse healthy microbe communities throughout life. Large polysaccharides are difficult for the small intestine to digest and the diverse species in the cecum and colon are more able to do this. The colon, also, has fewer molecules that can kill microbes and a much slower rate of flow. There are very few simple carbon molecules that are prevalent in the small intestine. These colon microbes do not need oxygen (anerobes) and degrade polysaccharides with the fermentation process (converting sugar to acids, gases or alcohol) This includes Bacteroidacae and Clostridia.
The cecum and large intestine are able to break down and digest plant fibers. Recently, it was found that eating fiber early in life is necessary to maintain diverse healthy microbe communities throughout life. Large polysaccharides are difficult for the small intestine to digest and the diverse species in the cecum and colon are more able to do this. The colon, also, has fewer molecules that can kill microbes and a much slower rate of flow. There are very few simple carbon molecules that are prevalent in the small intestine. These colon microbes do not need oxygen (anerobes) and degrade polysaccharides with the fermentation process (converting sugar to acids, gases or alcohol) This includes Bacteroidacae and Clostridia.
In the colon there are many creases, where the wall folds and lies over parts of itself. This creates specific niches and compartments in the inter fold areas. In the folds, there is a strikingly different environment than in the center of the lumen flow. The center region is sometimes called the digesta. The folded regions have a much greater amount of mucus (made by secretions from epithelial cells to create a special environment for friendly bacteria near the surface). These fight off dangerous types of bacteria. Mucus, also, has a lot of nutrients in it for those equipped to eat it.
Mucus and Crypt Environments
 Goblet cells exist in both the small and large intestine where they continuously secrete mucus to cover and protect the epithelial surface. The mucus in the small intestine is relatively small single layer of mucus. In the colon there are at least two distinct layers, one attached to the epithelium and the other loose and non-attached. The inner layer has almost no bacteria, since it is a barrier and has molecules that can kill bacteria. In the small intestine the epithelium secretes a great amount of local oxygen, which eliminates many of the more dangerous bacteria as well.
Goblet cells exist in both the small and large intestine where they continuously secrete mucus to cover and protect the epithelial surface. The mucus in the small intestine is relatively small single layer of mucus. In the colon there are at least two distinct layers, one attached to the epithelium and the other loose and non-attached. The inner layer has almost no bacteria, since it is a barrier and has molecules that can kill bacteria. In the small intestine the epithelium secretes a great amount of local oxygen, which eliminates many of the more dangerous bacteria as well.
The outer mucus is loose and parts of it break off and float in the center as islands of mucus. This can slow the flow in the center. The density increases in the ascending and transverse colon sections and then is decreased in the descending and sigmoid. This stimulates more bacteria that like mucus in the first part of the colon.
Unique Properties of Mucus
 Mucin (MUC2) forms links between molecules that make a mucus gel like. Wherever there is no mucin there are larger biofilms. Some bacteria are able to swim through the gel and eat molecules. The main way bacteria move on their own is with flagella, which triggers pattern receptors and immune responses (see posts on secretory machines and pattern receptors). At times, the very dangerous Salmonella is able to use its flagella to reach the epithelium.
Mucin (MUC2) forms links between molecules that make a mucus gel like. Wherever there is no mucin there are larger biofilms. Some bacteria are able to swim through the gel and eat molecules. The main way bacteria move on their own is with flagella, which triggers pattern receptors and immune responses (see posts on secretory machines and pattern receptors). At times, the very dangerous Salmonella is able to use its flagella to reach the epithelium.
Another strategy of dealing with mucin occurs with Shigella and E. coli, which make a protein (protease Pic) to eat the mucin. This stimulates much more secretion of mucin, which hurts the local friendly bacteria that have been there before. Proteobacteria, Firmicutes and Bacteroidetes make another enzyme, M60-like peptidases, that can do the same.
In the first part of the colon, a biofilm is formed that is consistent across many different animals including most mammals and amphibians. There are some special interactions that occur in this region to create the ubiquitous biofilm. A future post will describe the politics and geography of multiple microbes that create multiple three dimensional niches in biofilms.
In humans, the bacteria species of Actinobacteria and Proteobacteria are prominent in the mucosal region compared with the flowing lumen region digesta. Akkermansia muciniphilia are prominent bacteria that break down the mucus material in humans. Specific animal species have different unique microbe colonies for this task. Biopsies in humans show that there is wide variation of patches as small as one centimeter with different microbe species. Some of these patches are small and some are huge with saturating amounts of bacteria.
This same phenomenon of distinct patches exists in the feces as well with small and large communities juxtaposed in specific three dimensional regions. This is related to interactions of many different species, specific plant fibers available and other environmental factors. Therefore, the common use of relatively small feces samples to determine which microbes are present is inadequate, since it only captures several of these many different individual isolated patches. A previous post showed that interactions of communities often cause infections. Also, even in the one region that is in the sample, diversity is lost when doing a sample that is not maintained in its exact structure. Ordinary fecal samples make no attempt to maintain the three dimensional structure of the ingredients. Only with this rigorous research techniques can the real microbe patches be found.
There are some species that go all the way through the mucus and attach to the epithelium, especially in the colon crypts. These are very hard to study because washing samples disrupts the microfilms and leads to incorrect assumptions, such as that the surface does not have significant colonies. New techniques that preserve the mucus structure find many colonies in healthy human mucus and on the lining cells. These studies show special types that are able to use oxygen. These close colonies have immune protection or are ignored by the immune system. One example is B. fragilis that lives in crypts and uses immune alterations and special food. The old dogma that microbes are only attached during disease is clearly not correct.
Diet In Different Regions Affects Community Survival
 Fatty acids and small carbohydrates are absorbed in the small intestine. Therefore, in the colon, larger complex long chain carbohydrates (polysaccharides) are fermented by bacteria. Bacteroides spp. is well able to break down these polysaccharides. Microbes have many different mechanisms for this metabolism, but one of the prominent systems is called Sus (or starch utilization system), which uses enzymes attached to lipids on the surface of the bacteria. Sometimes these enzymes are secreted from the bacteria.
Fatty acids and small carbohydrates are absorbed in the small intestine. Therefore, in the colon, larger complex long chain carbohydrates (polysaccharides) are fermented by bacteria. Bacteroides spp. is well able to break down these polysaccharides. Microbes have many different mechanisms for this metabolism, but one of the prominent systems is called Sus (or starch utilization system), which uses enzymes attached to lipids on the surface of the bacteria. Sometimes these enzymes are secreted from the bacteria.
The term glycan and polysaccharide are interchangeable; they are both made from single sugars (monosaccharides) that are placed together through O-glycosidic linkages of monosaccharides. They can include glycoproteins, glycolipids, and proteoglycans. Microbes break glycans into oligosaccharides (smaller numbers of attached sugars). A transporter in the microbe then takes in these smaller sugar pieces. They break down both complex sugars from food and from the mucus glycan structure. The Sus system has the capacity to attack many different kinds of glycan. With it, B. Thetaiotaomicron can live in many different kinds of crypts.
 Complex large sugars from food control the center of the lumen digesta colonies. Meat and plant based diets attract very different microbe communities and they shift rapidly if the diet changes for a time. It is quite reversible for a short time. But, the mucosal communities are not affected rapidly and the longer-range diet controls permanent communities that are often hidden. In research where glycans are altered in the diet, such as adding fructan, fucose, and others found in human milk, microbe communities will change. But, some have the genes and enzymes to eat all of the different sugars. More efficient types take over when one particular sugar is emphasized. In animals that eat many different kinds of food, there are large dynamic changes in the lumen species.
Complex large sugars from food control the center of the lumen digesta colonies. Meat and plant based diets attract very different microbe communities and they shift rapidly if the diet changes for a time. It is quite reversible for a short time. But, the mucosal communities are not affected rapidly and the longer-range diet controls permanent communities that are often hidden. In research where glycans are altered in the diet, such as adding fructan, fucose, and others found in human milk, microbe communities will change. But, some have the genes and enzymes to eat all of the different sugars. More efficient types take over when one particular sugar is emphasized. In animals that eat many different kinds of food, there are large dynamic changes in the lumen species.
In the mucus regions there is stability with these rapid dietary changes in the lumen. Some species become the prominent indigenous communities and have access to the best niches and crypts. Mucin 2 (MUC2) is a very complex material with, at least, a hundred different surface structures in humans. These are linked by oxygen bonds to glycans that are different in each species—humans much different from mice. Types of mucin glycans are connected with favorite and special microbes that live near the epithelium. The microbes at the epithelium have to be able to resist the toxins sent by the lining cells.
 The microbe A. muciniphilia is very prominent in the mucus of humans and can eat the mucin. This means they can live without the need for other specific foods. They are very dominant during fasting. During inflammation, a different type of mucin eater is also prominent, B. Acidifaciens. The ability to eat primarily the mucin, rather than food is one of the major characteristics of species that stay for generations including being transferred from mother to children. In fact, the creation of the healthy mucus is arrived at by back and forth cooperation of the microbes, the epithelium and the immune system.
The microbe A. muciniphilia is very prominent in the mucus of humans and can eat the mucin. This means they can live without the need for other specific foods. They are very dominant during fasting. During inflammation, a different type of mucin eater is also prominent, B. Acidifaciens. The ability to eat primarily the mucin, rather than food is one of the major characteristics of species that stay for generations including being transferred from mother to children. In fact, the creation of the healthy mucus is arrived at by back and forth cooperation of the microbes, the epithelium and the immune system.
There are some nutrients that are only present briefly. Occasionally, particular bacteria can capitalize on this. Special factors (called colonization factors or CCFs) help B. fragilis and Bacteroides vulgatus become prominent for a while. There are special genes that are needed for this special niche. With these CCFs, these species are sometimes able to resist antibiotics and infections (gastroenteritis). There are some bacteria that have this narrow focus and can live in particular crypts for a long time. Some particular species love only an environment with a small amount of oxygen like the crypts.
Various Antimicrobials Are Produced By Epithelial Cells in Different Regions
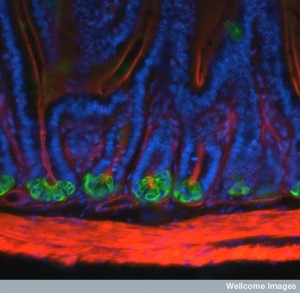 Paneth cells are at the base of the crypts and produce many molecules that can fight microbe species, especially in the beginning of the small intestine. A gradient of these antimicrobials is formed from this proximal secretion and many more species survive and exist near the end of the small intestine.
Paneth cells are at the base of the crypts and produce many molecules that can fight microbe species, especially in the beginning of the small intestine. A gradient of these antimicrobials is formed from this proximal secretion and many more species survive and exist near the end of the small intestine.
These are often peptides that have a positive charge, which affect the membranes of bacteria that have a negative charge. The outer membrane of bacteria is made up of a variety of molecules that include lipopolysaccharides (LPSs) that fight against these positively charged molecules from the Paneth cells. A section of the LPS has less than usual phosphate molecules, which helps resist the antimicrobials and increases colonization of both B. thetaiotaoemicron and Heliobacter pyroli.
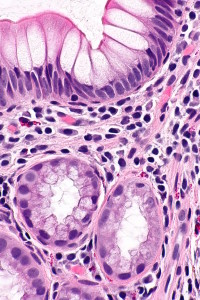
One important antimicrobial is called lectin REGIIIγ, which kills Gram-positive bacteria by damaging their outer layer of peptidoglycan. This particular molecule is necessary to avoid attack from a variety of dangerous species. The immune system contributes many other molecules to attack dangerous microbes (α and β-defensins) that attempt to come near the epithelium. Species such as Firmicutes and Bacteroides both have the ability to resist these molecules.
The epithelium also sends reactive oxygen molecules, which are difficult for most gut bacteria that are largely anaerobes. There is a gradient of more oxygen near the epithelium and little in the center of the lumen. The bacteria near the epithelium and mucus have special ways of dealing with oxygen. The bacteria B. fragilis is unusual in that it can handle small amounts of oxygen that exist in these special niches of the crypts.
Attaching to Epithelial Cells
 To attach to the epithelial cells bacteria have to go through the mucus. Some bacteria have special ways to attach to the epithelial cells. Helicobacter spp. is able to do this in the very acidic stomach as well as the small intestine by attaching to glycans. In the middle of the small intestine SFB (segmented filamentous bacteria) are able to grab onto the epithelium through an unknown mechanism. They use actin (filaments), which somehow makes a hole in the epithelial cells. SFB also influence the immune system allowing them close to the lining.
To attach to the epithelial cells bacteria have to go through the mucus. Some bacteria have special ways to attach to the epithelial cells. Helicobacter spp. is able to do this in the very acidic stomach as well as the small intestine by attaching to glycans. In the middle of the small intestine SFB (segmented filamentous bacteria) are able to grab onto the epithelium through an unknown mechanism. They use actin (filaments), which somehow makes a hole in the epithelial cells. SFB also influence the immune system allowing them close to the lining.
One mechanism of attachment used by dangerous disease causing microbes has been discovered. These use proteins on the membrane including lectins, adhesins and large appendages called fimbriae or attachment pili. (see post on secretory machines). Vibrio cholera uses pili with toxins. They have special proteins on the outer membrane. If they cannot attach, they are not dangerous. E. coli has many different kinds of lectins, with many different kinds of sugars that are useful to attach to epithelial cells. Listeria uses multiple steps—first an adhesion protein, then actin structures that open the epithelial cell.
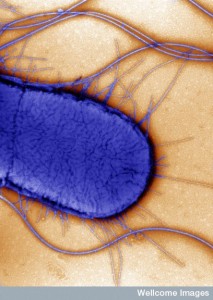 But, the regular permanent residents are not well known yet. In fact, for a long time it was thought that there weren’t any there. Friendly, permanent species use up the binding sites that keep away the dangerous types. Lactobacillaeceae have pili, membrane proteins that adhere to epithelial cells as well as the mucus and extracellular matrix. Mucus binding proteins (MUBs) are critical for lactobacillaceae to help stabilize the gut microbes. There are many different factors that allow microbes to form groups and to grab onto the epithelial cells that are then protective. These form friendly biofilms that are protective
But, the regular permanent residents are not well known yet. In fact, for a long time it was thought that there weren’t any there. Friendly, permanent species use up the binding sites that keep away the dangerous types. Lactobacillaeceae have pili, membrane proteins that adhere to epithelial cells as well as the mucus and extracellular matrix. Mucus binding proteins (MUBs) are critical for lactobacillaceae to help stabilize the gut microbes. There are many different factors that allow microbes to form groups and to grab onto the epithelial cells that are then protective. These form friendly biofilms that are protective
Both the friendly and the dangerous types, in fact, appear to use similar techniques.
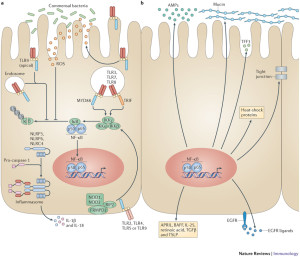
Conversations with Immune Cells Are Critical
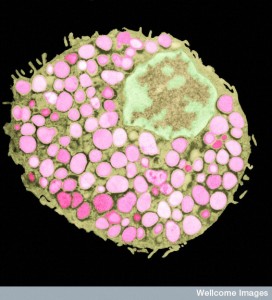 Friendly bacteria near the epithelium are tolerated. A particular immunoglobulin sIgA coats many of the microbes that are being protected. Some are especially coated such as SFB and Helicobacter spp. This allows friendly biofilms as a barrier to dangerous types. The immunoglobulin helps the co existence of the microbes, the epithelium and immune cells. There are special antibodies produced for the sugar molecules in the capsule of the friendly microbes. In fact, it is not yet clear how the immune system is able to tell the difference between some friendly and unfriendly microbes. But, they do.
Friendly bacteria near the epithelium are tolerated. A particular immunoglobulin sIgA coats many of the microbes that are being protected. Some are especially coated such as SFB and Helicobacter spp. This allows friendly biofilms as a barrier to dangerous types. The immunoglobulin helps the co existence of the microbes, the epithelium and immune cells. There are special antibodies produced for the sugar molecules in the capsule of the friendly microbes. In fact, it is not yet clear how the immune system is able to tell the difference between some friendly and unfriendly microbes. But, they do.
In some situations, the microbes manipulate the immune system (discussed in other posts). B. fragilis alters the immune system. The capsule polysaccharide signals with a dendritic immune cell to create interleukin 10 (IL-10) from T cells. These cells protect the microbe. This type of mechanism occurs with other microbes, as well, that decrease the amount of cytokines promoting inflammation and lowering B lymphocytes. SFB triggers a special type of T cell (Th17), which specially protects them. In fact, there are many different versions of this type of modulation of immune cells. Some involve interactions with as many as eight different microbe species at the same time.
There are general signals put out by epithelial cells such as specific fatty acids that trigger regulatory T cells that help particular types of microbes. Mucus, also, stops inflammation, inhibiting dendritic cells and cytokines. Mucin eating microbes are involved in this process. Many dangerous microbes have been known to trick immune cells to stop inflammation. What is very confusing is that particular factors that have been associated with dangerous microbes, such as pili, capsular sugars, eating mucus, adherence to epithelial cells, and altering immune functions, are also used by the friendly types in the same way
It does appear that this is an intelligent collaboration of the three types of cells—many different microbes, many different immune cells and several different epithelial cells.
Gut Geo Politics
 After birth, there is a dominant species that gradually changes into many different species. This specialization in many different niches occurs starting in the beginning of the small intestine and travels downward along with the one-way flow. With disease, changes in behavior and antibiotics eliminate many of the species in the moving center, not in the special hidden niches.
After birth, there is a dominant species that gradually changes into many different species. This specialization in many different niches occurs starting in the beginning of the small intestine and travels downward along with the one-way flow. With disease, changes in behavior and antibiotics eliminate many of the species in the moving center, not in the special hidden niches.

These niches, along with the well-protected appendix, create a long lasting reservoir that replenishes the friendly species. The appendix has a unique varied population of immune cells as well as a diverse set of microbes in niches. It is involved in the education of lymphocytes for the modulation of the reactions to friendly species. Despite dominant older views, the appendix, in fact, evolved two separate times and exists in a wide range of species. It hardly appears to be an accident. The crypts, the niches near the mucus and the appendix are very involved in the regulation and stabilization of the immune system and the diversity of species, especially after disasters that wipe out most of the lumen species.
There are strong associations with alterations in the status quo of microbe species and immune cells to many diseases in the gut and elsewhere—metabolic, immune and brain.
Inflammatory bowel disease (IBD) has less Clostridia along with fewer species in general. In childhood, also, there is less diversity. Unfortunately, studies of many diseases are poor because they use fecal samples, which is not a good way to find out species, instead of the more difficult biopsies of the inner lining.
 Inflammatory bowel disease studies of fecal material show a wide range of species with less diversity than normal. Those related to the disease are from near the mucus and epithelial cells and are coated with the immunoglobulin sIgA. Biopsies show more bacteria than normal in the mucus, less diversity and less Clostridium spp and increased amounts of dangerous E. coli. The studies of fecal samples didn’t show these findings.
Inflammatory bowel disease studies of fecal material show a wide range of species with less diversity than normal. Those related to the disease are from near the mucus and epithelial cells and are coated with the immunoglobulin sIgA. Biopsies show more bacteria than normal in the mucus, less diversity and less Clostridium spp and increased amounts of dangerous E. coli. The studies of fecal samples didn’t show these findings.
With serious damage to the liver (cirrhosis) there is often mental confusion (encephalopathy). Toxins made by microbes are not destroyed by the liver and go into the brain. Fecal material didn’t show much difference, but the mucus did. There was less Roseburia spp. and much more Enterococcus, Veillonella, Megasphaera and others. These species found in the mucus and near epithelial cells appear to be related to more inflammation and less cognition. Again, it is the mucus and not the fecal stream that is important.
Too Much Cooperation is Not Stable
 A counter intuitive recent finding shows that too much cooperation between species causes perpetual positive feedback, which has extreme effects. This destroys competing populations, which, surprisingly are needed to increase stability of both. By increasing the strength of both communities too much, an unrealistic situation is created of too much growth that can’t be sustained.
A counter intuitive recent finding shows that too much cooperation between species causes perpetual positive feedback, which has extreme effects. This destroys competing populations, which, surprisingly are needed to increase stability of both. By increasing the strength of both communities too much, an unrealistic situation is created of too much growth that can’t be sustained.
Because too much positive feedback causes a collapse of both, the immune system stops this type of positive feedback. The host separates cooperating species. The host also provides different sources of food (carbon) for each. Cooperation is not common and only is 10% of interactions.
Clearly, a certain amount of cooperation is necessary. But, also, separation and competition lead to long-term stability. This research shows that we have a lot to learn about this immensely complex set of relationships between these communities.
Microbe Geo Politics in the Gut
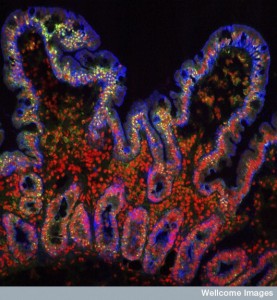 Over the vast expanse of the gastro intestinal tract, there are thousands of patches, villages and some diverse large cities of microbes of many types. They can live in the fast moving river in the middle of the small intestine lumen. Or they can live in the deep crypts below the protective mucus layer. Some brave the stark acid conditions of the stomach. Some can eat the mucus that kills most species. Some of the communities receive help and blessings from the epithelial lining cells to live close by. Some receive communication and help from the many different immune cells. The constant chatter between the epithelial and immune cells is extremely complex. With this back and forth communication, these many different diverse communities are surprisingly stable over time.
Over the vast expanse of the gastro intestinal tract, there are thousands of patches, villages and some diverse large cities of microbes of many types. They can live in the fast moving river in the middle of the small intestine lumen. Or they can live in the deep crypts below the protective mucus layer. Some brave the stark acid conditions of the stomach. Some can eat the mucus that kills most species. Some of the communities receive help and blessings from the epithelial lining cells to live close by. Some receive communication and help from the many different immune cells. The constant chatter between the epithelial and immune cells is extremely complex. With this back and forth communication, these many different diverse communities are surprisingly stable over time.
Recently, it was learned that the three dimensional structure of feces is produced by the same complex interactions of many communities. Only by studying intact feces are we learning what is really there and what the dynamics are. Also, biopsies all along the lining have shown that it is not sterile at all, but has many chosen friendly communities in each region. And, occasionally, some not so friendly. Somehow, all of this depends upon intelligent chatter of thousands of cells.
The next post will discuss the complex three-dimensional structures of biofilms, which, surprisingly, are, also, made up of multiple interacting species.