 While all cells are complex, the neuron is vastly complex. It is hard to understand how so many different mechanisms all through the cell can be correlated with mental events. Previous posts have clarified very complex processes in both the axons and dendrites. One post noted how materials are sorted, tagged and transported along the axon to where they are needed. Different motors and adaptors are required for the movement of varied cargoes. Another post showed how little is yet known about the extremely rapid complex process of producing, maintaining, secreting and reloading vesicles with neurotransmitters at the synapse. Another post showed that in the dendrite, signaling between the thousands of sections in the arbor is much more complex than previously understood.
While all cells are complex, the neuron is vastly complex. It is hard to understand how so many different mechanisms all through the cell can be correlated with mental events. Previous posts have clarified very complex processes in both the axons and dendrites. One post noted how materials are sorted, tagged and transported along the axon to where they are needed. Different motors and adaptors are required for the movement of varied cargoes. Another post showed how little is yet known about the extremely rapid complex process of producing, maintaining, secreting and reloading vesicles with neurotransmitters at the synapse. Another post showed that in the dendrite, signaling between the thousands of sections in the arbor is much more complex than previously understood.
Recent research has tried to tackle the way that the neuron is able to keep the environments of the axon and the dendrite so totally different. Research has begun to clarify how axons and dendrites develop, mature and change with neuroplasticity. Neurons are called polar cells because the functions of the dendrites and axons are so unique. Much of this is based on the different scaffolding and proteins that are enmeshed in the various membranes to provide the electro chemical basis for receiving and sending signals.
 All of these are produced near dendrites and the beginning of the axon. But, two completely different sets are then sorted, tagged and transported. In fact, this process is extraordinarily complex involving different types of transport systems, motors, adaptors, vesicles and proteins. In addition, all of this has be regulated. It is only recently, that research has tackled possible mechanisms to maintain the neuronal polarity.
All of these are produced near dendrites and the beginning of the axon. But, two completely different sets are then sorted, tagged and transported. In fact, this process is extraordinarily complex involving different types of transport systems, motors, adaptors, vesicles and proteins. In addition, all of this has be regulated. It is only recently, that research has tackled possible mechanisms to maintain the neuronal polarity.
One of the major issues is tracking the specific vesicles that are needed to transport each specific component to its exact polar destination. Protein regulation is needed to produce these vesicles—budding from a membrane, transport and then fusion at the destination. Currently, we don’t even know how many vesicles types are needed and for each there is needed specific different coat proteins and adaptors to hold the cargo and varied signaling.
 Each needs motors, adaptors, tethers and SNARE molecules that allow the vesicle to fuse and deliver its cargo. How are the vesicles directed in these two different. For the dendrite they are sent to the dendrite membrane. In the axon there are many long distance transport systems. How are the two systems coordinated so that they don’t go in the wrong direction or are redirected when they do? How are all of these parts sorted, tagged and transported?
Each needs motors, adaptors, tethers and SNARE molecules that allow the vesicle to fuse and deliver its cargo. How are the vesicles directed in these two different. For the dendrite they are sent to the dendrite membrane. In the axon there are many long distance transport systems. How are the two systems coordinated so that they don’t go in the wrong direction or are redirected when they do? How are all of these parts sorted, tagged and transported?
Polarized Neurons
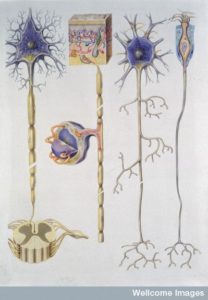
 Neurons have many dendrites and one long axon. The axon can break at its tip into some smaller branches as well, but nothing like the possibly 10,000 dendrites with hugely complex branching arbors. The branching arbors constantly change and have elaborate signaling between dendrite heads, close and far, and in different geometrical configurations. Axons and dendrites are very different compartments of the cell and have very particular proteins in the membranes, such as many channels and varied receptors with different functions. The scaffolding is different. In fact, all molecules in both regions need extremely accurate placement.
Neurons have many dendrites and one long axon. The axon can break at its tip into some smaller branches as well, but nothing like the possibly 10,000 dendrites with hugely complex branching arbors. The branching arbors constantly change and have elaborate signaling between dendrite heads, close and far, and in different geometrical configurations. Axons and dendrites are very different compartments of the cell and have very particular proteins in the membranes, such as many channels and varied receptors with different functions. The scaffolding is different. In fact, all molecules in both regions need extremely accurate placement.
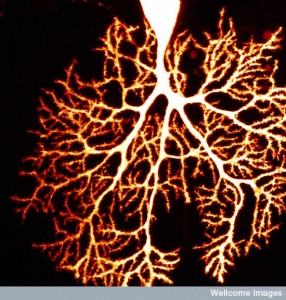 have very different structures in the two regions—active zones for staging of neurotransmitter vesicles versus post synaptic densities, spines and heads. But, almost all of the basic production of vital molecules occurs in the same endoplasmic reticulum and Golgi in the cell body. In recent years, research into how vital proteins, in particular, are separated and transported. Most of the research has been in the hippocampus and the cortex.
have very different structures in the two regions—active zones for staging of neurotransmitter vesicles versus post synaptic densities, spines and heads. But, almost all of the basic production of vital molecules occurs in the same endoplasmic reticulum and Golgi in the cell body. In recent years, research into how vital proteins, in particular, are separated and transported. Most of the research has been in the hippocampus and the cortex.
New proteins are placed in vesicles that bud from the Golgi network and then are transported along microtubules to the axon or the dendrite. The proteins are brought to the membrane and then extruded from the vesicle into the membrane. A large system of vesicles called endosomes are part of the cellular transport system. This same process occurs in all cells, but is more extreme in the huge, geometrically complex neurons. Problems in transport of these vital proteins produces many brain diseases.
There are another large number of proteins that produce the transport, itself. The first budding of vesicles is regulated by coat proteins, transported from the cell body to the membrane. Each protein must have particular cargo adaptors. There are others that make the membrane curved in particular ways. Proteins are sorted and attached to large other proteins sitting across the membrane. More complex molecules have sugar attachments to proteins—glycoproteins. These have unique adaptors and receptors as well.
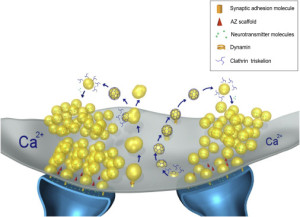 As well as budding, the fusion of the vesicles has another large set of proteins that link the vesicles to membranes. A large set of proteins are tethering proteins that make first contact. Others called SNARE’s (discussed in detail in relation to neurotransmitter vesicle release) fuse the vesicle and membrane.
As well as budding, the fusion of the vesicles has another large set of proteins that link the vesicles to membranes. A large set of proteins are tethering proteins that make first contact. Others called SNARE’s (discussed in detail in relation to neurotransmitter vesicle release) fuse the vesicle and membrane.
The transport of the vesicle along microtubules is another complex system using many different motors—kinesins and dyneins. These move in opposite directions (see post on neuronal transport). These each have multiple adaptor molecules for each type of cargoes. Motor adaptors comprise a large set of different kinds of proteins, not one family. All of this regulated by enzymes that use energy called RABs and ARF.
The microtubules have different electric charges in different directions and these direct some of the traffic to dendrites or axons. While the proteins can be transported back and forth dendrite, material must not go into the axon. Some axonal material goes into the dendrite but not the opposite. Some material is sent into the dendrite where extrusion from the vesicle is rapid and those not put in place are prevented from being released and then lose their polarity.
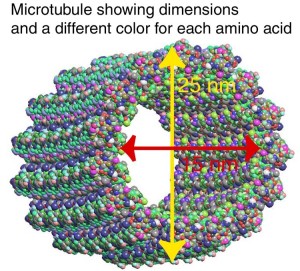 The situation is more complex in the axon which is much bigger with more back and forth transport. Complicating the situation is that the endoplasmic reticulum and Golgi extend someone into the dendrite tree. There are two to four times as many vesicles going into the axon. One particular pathway sends proteins from Golgi only to the axon and not the dendrite. To make it more complex, most proteins are not just sent to the axon, but to very particular sections of the axon such as the node of Ranvier in the myelin structure or the compartments of the synapse.
The situation is more complex in the axon which is much bigger with more back and forth transport. Complicating the situation is that the endoplasmic reticulum and Golgi extend someone into the dendrite tree. There are two to four times as many vesicles going into the axon. One particular pathway sends proteins from Golgi only to the axon and not the dendrite. To make it more complex, most proteins are not just sent to the axon, but to very particular sections of the axon such as the node of Ranvier in the myelin structure or the compartments of the synapse.
Once in a region, these proteins must be prevented from moving along the membrane to another region. Special scaffolding proteins sit just under the membrane and hold the proteins in place. Specific channels, such as the sodium channels used in the nodes of Ranvier, are maintained by special actin networks. Those proteins that are not bound in this way cannot get by a barrier in the axon initial segment.
Sorting in Dendrites
 As proteins are finalized in the Golgi, they are sorted between those going to dendrites and the axon. They are placed in specific vesicles that are transported on microtubule highways. Those to the dendrites can go in either direction, but never go to the axon. Axon vesicles can go to the dendrite but are rerouted to the axon.
As proteins are finalized in the Golgi, they are sorted between those going to dendrites and the axon. They are placed in specific vesicles that are transported on microtubule highways. Those to the dendrites can go in either direction, but never go to the axon. Axon vesicles can go to the dendrite but are rerouted to the axon.
First, proteins must be in the particular vesicles. Special sorting signals between adaptors and cargo proteins place them in the proper vesicles. Sorting signals were first discovered in the dendrite proteins. They are part of proteins that cross the membrane in the side inside the cell. There are five specific large molecular complexes called adaptor protein complexes (AP1, 2, 3, 4, 5).
These have four subunits, with different versions in different sizes. The many different types work at different locations in the cell. Some bind to the cell’s membrane, others to the Golgi and others to the endosome vesicles. These are sorted through specific signals that are three dimensional structural sequences on the proteins that bind to other surfaces. Some of these signals are in complex lipo-proteins (large proteins that have fat molecules attached (see post).
Many Different Proteins
Proteins are polarized as well, meaning they have different electrical charge in different parts of the molecule. The simplest version of a polarized molecule are fatty acids that have one part that likes water and other that likes fat. This allows two such molecules to create membranes that keep water out by having the water hating part of both molecules inside the membrane and the water loving part on the outside of the membrane.
Polarized proteins are more complex and have a very large amount of shapes and possibilities. These different polarized proteins exist in clusters in specific regions of the neuron—such as the multiple different nodes related to myelin on the axon and the synapse. The research on these different proteins is just beginning. But, factors include diffusion along the membrane until a certain region is found and different scaffolding in each region. But, in the long axon with many barriers to diffusion, specific proteins must be delivered exactly by microtubule transport. As of now the exact polarization of the many proteins and the ways this translates into exact transport are not known.
Some of the proteins regulating this process:
- Sorting: Adaptors to Clathrin; Adaptors put transmembrane cargo proteins in Golgi into vesicle buds; Sorting nexins regulate plasma membrane, endosomes, and Golgi
- Transport: kinesins, kinesin adaptors, dyneins, dynein adaptors. Dynactin complex connectings both motors.
- Fusion: tethering factors initial interaction; SNARE forces membrane mixing and fusion;
- Further Stages: ARF (ADP ribosylation factor) 29 different GTP enzymes; RAB (ras – related in brain) GPT enzymes with 60 different types;
Axon sorting
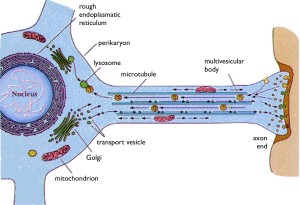 There is more known about dendrite sorting, because it is a much smaller space closely connected to the cell body. Along the huge axon, the exact mechanism is not known. There have been specific sequences in some of the proteins that have been identified as sequences in this transport. One line of research relates to the vital potassium channels. Please see the post on the vast complexity of the 100 different types of potassium channels.
There is more known about dendrite sorting, because it is a much smaller space closely connected to the cell body. Along the huge axon, the exact mechanism is not known. There have been specific sequences in some of the proteins that have been identified as sequences in this transport. One line of research relates to the vital potassium channels. Please see the post on the vast complexity of the 100 different types of potassium channels.
K1 seems to be related to the transport of these specific proteins in the hippocampus. They are mostly on the axon initial segment and paranodes (near the node of Ranvier – see post on myelin). Specific polarization is found where there is no myelin. But, it is not yet clear whether these relate to budding process or delivery process. The first sorting receptors in the axon have just been discovered in a worm.
Start of Microtubule Motor Proteins
 Once vesicles are sorted and tagged, then they are sent on specific types of transports. A previous post described the vast complexity of this transport along the extremely long axon. As a matter of scale, one axon from the spinal cord to the foot is three feet long and compared to the size of the cellular components is equivalent to walking along the wall of China. The complexity described included the many specific motors and adaptors needed for each type of transport.
Once vesicles are sorted and tagged, then they are sent on specific types of transports. A previous post described the vast complexity of this transport along the extremely long axon. As a matter of scale, one axon from the spinal cord to the foot is three feet long and compared to the size of the cellular components is equivalent to walking along the wall of China. The complexity described included the many specific motors and adaptors needed for each type of transport.
Microtubules are also different in the axon and dendrite. Almost all of the axon microtubules have their negative ends in the direction of the cell body and the plus end in the direction of the synapse. In dendrites they can be either direction. Near the end of the dendrite most are oriented with leading negative end.
Varied Motors For Each Purpose
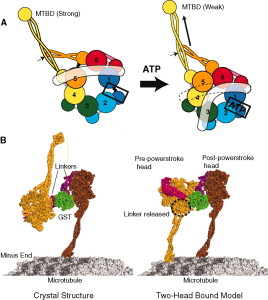 In axons, kinesin moves forward to the synapse and dynein backwards. But, also all of the associated proteins are different and the proteins have unique tags and attachments to the subunits called tubulin. These unique modifications create chemical changes so that different parts of the microtubule have unique properties
In axons, kinesin moves forward to the synapse and dynein backwards. But, also all of the associated proteins are different and the proteins have unique tags and attachments to the subunits called tubulin. These unique modifications create chemical changes so that different parts of the microtubule have unique properties
There are more than 20 kinesin motors with different properties in the plus direction on microtubules. Dynein has a heavy chain subunit and many secondary subunits. Kinesins walk for a short way carrying a vesicle and then reconnect with the microtubule and continue walking. Specific different kinesin motors are used in the axon and dendrite. Some uniquely carry mitochondria. In the dendrite several carry receptors and protein channels for the membrane in the dendrite.
Unique Initial Segment

A previous post noted the unique environment of the initial segment of the axon in determining transport along the axon. But only recently has its role in polarity been found. In the initial segment there is little movement of proteins in the membrane itself. They are anchored as are complex lipid molecules bound to proteins. Even those not bound to the actin scaffold cannot move much because of the actin structures. Therefore, in the initial segment active localization is maintained by sorting, tagging and transport.
Chemical Differences
 Tubulin is the molecule that builds microtubules. Tagging and alterations of this protein produces different types of microtubules that are only now being discovered. Modifications of this vital building block protein include acetyl placed on a lysine that is in the center of the round tubule. Another alterations includes glutamine near one end (carboxyl terminus) and removal of a tyrosine. These changes appear to be relevant to the particular motors used. Specific motors walk better with these changes. With these changes occurring in axon tips, there are more motors attracted to this location.
Tubulin is the molecule that builds microtubules. Tagging and alterations of this protein produces different types of microtubules that are only now being discovered. Modifications of this vital building block protein include acetyl placed on a lysine that is in the center of the round tubule. Another alterations includes glutamine near one end (carboxyl terminus) and removal of a tyrosine. These changes appear to be relevant to the particular motors used. Specific motors walk better with these changes. With these changes occurring in axon tips, there are more motors attracted to this location.
While some of the axon motors can go into the dendrite, factors move them toward the axon. There are many more plus end microtubules in the axon. Also the microtubules heading for the axon have alterations that attract the specific motors and adaptors.
Vesicles tagged for the dendrite are stopped at the axon initial segment. Some are sent back and others fuse with the local membrane. It is not really clear how this occurs but there is new data supporting some theories. With mixed polarity of tubules in the dendrite, dynein could go in two directions. Dynein would not bring them into the axon. However, it is known that some kinesins work in the dendrite.
Another set of data shows that myosin motors (see post) work in the dendrite. Vesicles transported int the initial segment come up against dense actin scaffolding. Myosin is known to interact and work with actin and pulls them part of the way back toward the cell body and dendrite.
Selective Vesicle Fusion
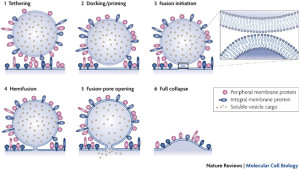 Much research has been devoted to the complex fusion of vesicles with the membrane in the synapse. Despite a massive amount of work, these mechanisms are still in question (see post on synapse vesicles). Much less work has been done related to how these vesicles fuse when bringing receptors, ion channels and other materials. The complexity of tethering adaptors and SNARE mechanisms have been mentioned above. The many proteins and types of SNARE are just now under investigation.
Much research has been devoted to the complex fusion of vesicles with the membrane in the synapse. Despite a massive amount of work, these mechanisms are still in question (see post on synapse vesicles). Much less work has been done related to how these vesicles fuse when bringing receptors, ion channels and other materials. The complexity of tethering adaptors and SNARE mechanisms have been mentioned above. The many proteins and types of SNARE are just now under investigation.
How Does A Neuron Stay Polarized
 Transport of material along the huge axon has been described as a vastly complex process. Signaling between the fantastic number of dendrites is another immense process.
Transport of material along the huge axon has been described as a vastly complex process. Signaling between the fantastic number of dendrites is another immense process.
Now, research shows that maintaining the polarity of the neuron—the enormous difference between the various compartments of the axon and dendrites—is a massive job. It involves many special molecules for sorting, transport and fusion to the membranes. Each of these jobs involves special adaptors, vesicles, scaffolding, motors, and fusion mechanisms. Each needs unique proteins and lipoproteins.
With no obvious direction, these fantastic simultaneous processes occur in sync with mental events.